
Marvel I. Adedeji
Department of Veterinary Public Health and Preventive Medicine, University of Ibadan, Ibadan, Nigeria
Victoria O. Adetunji
Department of Veterinary Public Health and Preventive Medicine, University of Ibadan, Ibadan, Nigeria
LiveDNA: 234.1198
Research Journal of Microbiology
Year: 2020 | Volume: 15 | Issue: 3 | Page No.: 90-97







ABSTRACT
Background and Objective: Ginger is well known for its medicinal and preservative values but its use for inhibition and dispersion of E. coli O157:H7 biofilm has not been sufficiently explored. This study was therefore designed to evaluate the biofilm inhibition and dispersion activities of Zingiber officinale crude extract and fractions on biofilms of E. coli O157:H7 isolated from dispensed powdered milk in Ibadan, South-West Nigeria. Materials and Methods: Ginger root samples were obtained, identified (UIH-22558), extracted, fractionated using four solvents (hexane, chloroform, ethyl acetate and ethanol) and concentrated. Three E. coli O157:H7 isolates were obtained from dispensed powdered milk and a reference strain (E. coli ATCC 35218) was obtained from the Department of Microbiology laboratory, University of Ibadan. Modified crystal violet assay was employed to develop biofilm and to test the effect of plant extracts on biofilm formation and biofilm dispersal. Results: All the isolates developed biofilm (0.08±0.07) and biofilm dispersion was exhibited with the ethanolic (0.04±0.05) and crude extract fractions (0.05±0.06) having the highest effect, while chloroform fraction had little or no effect at 50 mg mL–1 concentration. Conclusion: Ginger (Zingiber officinale) showed potential use for dispersion of already formed biofilm by E. coli O157:H7 strains, hence, can be used in food processing plants, surfaces and industries to combat biofilm-forming organisms, disperse their biofilms and enhance food safety.
PDF Abstract XML References Citation
Received: December 02, 2019;
Accepted: February 11, 2020;
Published: June 15, 2020
How to cite this article
Marvel I. Adedeji and Victoria O. Adetunji, 2020. Antibiofilm Activities of Ginger (Zingiber officinale) on E. coli O157:H7 Isolates from Powdered Milk in Ibadan, Nigeria. Research Journal of Microbiology, 15: 90-97.
URL: https://scialert.net/abstract/?doi=jm.2020.90.97
URL: https://scialert.net/abstract/?doi=jm.2020.90.97
INTRODUCTION
Biofilm is a population of microbial cells that are enclosed within a matrix made of primarily polysaccharide material which can stick to surfaces1,2 and they can grow in natural, clinical and industrial surfaces. Studies have shown that biofilms are difficult to remove from food contact surfaces and equipment by normal cleaning, thus posing a food safety risk3. Bacteria protected within biofilm exopolysaccharides are up to 1,000 times more resistant to antibiotics than planktonic cells (free-floating), which generates serious consequences for therapy and severely complicates treatment options. An estimated 75% of bacterial infections involve biofilms that are protected by an extracellular matrix, hence, the ineffectiveness of antibiotic treatment in the biofilm diseases which causes serious problems in the eradication of infections4. Bacterial attachment, colonization and biofilm formation on food and food contact surfaces can serve as a source for bio-transfer and cross-contamination of the finished product, leading to health issues such as food poisoning and increased product spoilage5,6. The buildup of biofilms can affect the heat flow across a surface and increase surface corrosion and frictional resistance of fluids leading to loss of energy in a system, substantial economic losses in engineering systems7 and overall loss of products.
Escherichia coli (E. coli) is a Gram-negative, rod-shaped bacterium commonly found in the lower intestine of warm-blooded organisms (endotherms)8. E. coli O157:H7 is the most important entero-hemorrhagic E. coli (EHEC) serotype in relation to public health9. E. coli O157:H7 is well known as a major Shiga toxin-producing food-borne pathogen that has a low infectious dose and causes hemorrhagic colitis, hemolytic uraemic syndrome and even death10,11. Interaction of E. coli O157:H7 with intestinal epithelial cells is the first step in the colonization of its host cells and it possesses several effectors that allow it to adhere to intestinal epithelial cells10,12. Following initial adhesion to, bacteria begin to grow as a monolayer on the surface to form microcolonies that can consequently develop and form a biofilm. Hence, bacterial adhesion is an important virulence factor that plays an important role in gastrointestinal infectious disease13. E. coli strains are harmless, but some serotypes can cause serious food poisoning in humans and are occasionally responsible for product recalls due to food contamination14. Earlier reports have shown that the thickness of E. coli biofilm may be of hundreds of microns and posing a difficulty in treatment with antibiotics due to presence of exo-polymers and reported that a strain of E. coli O157:H7 produced significantly more biofilm than the other on a particular surface type15,16. Depending on the species involved, microcolonies may be composed of 10-25% cells and 75-90% extracellular polymeric substances (EPS) matrix17. The presence of pathogenic bacteria in the biofilm matrix increases their virulence and resistance to disinfectants and antibiotics18.
Studies have shown that two different anti-biofilm mechanisms are able to modulate biofilm formation: inhibition of bacterial surface attachment and destabilization/disruption of mature biofilms irreversibly attached. Many of the existing anti-biofilm agents are non-biocidal, but some bactericidal molecules could be considered as anti-biofilm agents as they are still active against mature biofilms protected by their architecture. Bactericidal anti-biofilm agents should be very specifically targeted otherwise their use could impair the composition of established ecosystems and damage beneficial microbiota. Nevertheless, anti-biofilm strategies represent interesting approaches for medical biotechnology as attested by a large number of recent publications7. Anti-biofilm agents fall into two large groups, synthetic and natural. Plant extracts could be considered as promising alternative non-antibiotic strategy aimed to reduce the production of virulence factors without affecting bacterial growth, which may be less prone to develop drug resistance19.
Ginger (Zingiber officinale) is a flowering plant whose rhizome has been widely used as a culinary or medicinal herb for thousands of years20. Ginger rhizome possesses two constituents; the essential oils, which give the aroma and 6-, 8- and 10- gingerols, which are considered as main pungent principles21. During thermal processing or storage, the gingerols may be modified to a series of homologous compounds of shogaols such as 6-, 8- and 10-shogaol22,23. Ginger can be used in a variety of forms. It has strong medicinal, preservative and anti-oxidative properties and thus contributes to the overall safety and preservation of foods. Recent studies have shown that ginger has antibacterial activity against Staphylococcus aureus and Streptococcus pyogenes that is higher than that of commercially available antibiotics24,25. Antibacterial alkylated gingerols from ethanol and n-hexane extracts of ginger have been isolated by Park2. Essential oils (ethanolic extract) of ginger have been studied for their antimicrobial activity against different bacteria including E. coli, S. typhi26,27. Moreover, essential oils of ginger have been used for preventing biofilm formation by Pseudomonas aeruginosa PA1428 Staphylococcus aureus29 and Candida albicans30, E. coli ATCC 25922, but did not show anti-biofilm effect against S. Typhimurium8. Other than the antibacterial properties that had been displayed, ginger is also known to have anti-tumorigenic, anti-inflammatory, anti- apoptotic and etc. characteristics31. However, the biofilm inhibition and biofilm dispersion effects of ginger on E. coli O157:H7 have not been studied.
There are strong contentions about the safety of conventional chemical antimicrobials and anti-biofilms because of the problem of resistance and other effects such as teratogenicity and carcinogenicity, hence, the rise in demand for natural substances which possess good antimicrobial and anti-biofilm effects32. The exploration of naturally occurring antimicrobials for food preservation is receiving increased attention due to consumer awareness of natural food products and a growing concern of microbial resistance towards conventional preservatives11.
Quorum Sensing (QS) is a mechanism that controls coordinated bacterial behaviors in response to the density of bacterial cells and is tightly linked to bacterial biofilm formation33,34 as well as to the production of virulence factors35-37 hence, it is the most extensively studied approach.
Many spices and herbs possess and exert antimicrobial activity due to their essential oil fractions and ginger Zingiber officinale is one of such. Therefore, this paper assesses the biofilm inhibition and dispersion activities of Zingiber officinale crude extracts and concentrated fractions of four solvents (hexane, chloroform, ethyl acetate and ethanol) on E. coli O157:H7 isolates from dispensed powdered milk.
MATERIALS AND METHODS
Study area: The geographical location for this study is, Oyo State in South Western Nigeria. Fresh samples of ginger (Zingiber officinale) were purchased at Ojoo market in Ibadan metropolis, Nigeria. Identification of the samples was done at the Department of Botany, University of Ibadan, Nigeria. The milk samples were also obtained from seven different markets in Ibadan, Nigeria. The study was carried out at the Meat and Milk Hygiene Laboratory, University of Ibadan, Nigeria. Plant extraction was done at the Pharmaceutical Chemistry Laboratory, Faculty of Pharmacy, University of Ibadan, Nigeria. Research work spanned through a period of ten months (November 2016-August 2017).
Plant preparation: Ginger root samples were obtained from the Ojoo market and identified at the herbarium of the Department of Botany, University of Ibadan (UIH-22558). The plant was extracted by cold maceration, solvent-solvent fractionation was done using four solvents (hexane, chloroform, ethyl acetate and ethanol) based on the degree of polarity and resulting fractions were concentrated using a rotary evaporator.
Sample preparation and pre-enrichment: The dispensed powdered milk samples were homogenized and pre-enriched using buffered peptone water and incubated at 37°C for 24 hrs. Selective enrichment was done, using modified tryptone soy broth which was dissolved in distilled water, sterilized by autoclaving and left to cool to 45°C in a water bath before the addition of novobiocin which was meant to suppress the growth of other Gram-negative bacteria and also serve as a medium of pre-enrichment. To the homogenized milk sample, 9 mL of the mTSB was added and incubated at 37°C for 24 hrs.
Isolation and identification of E. coli O157:H7: Three E. coli O157:H7 isolates were obtained from pre-enriched samples using standard cultural, biochemical and culture plating methods and a reference strain (E. coli ATCC 35218) was obtained from microbiology laboratory. Sorbitol MacConkey agar was used as the selective differential solid medium, 0.1 mL of the milk broth culture was inoculated using surface plating method and incubated at 37°C for 18-24 hrs. Presumptive colonies were subcultured on freshly prepared SMAC. Clear, distinct colonies were inoculated onto nutrient agar slants and stored in the refrigerator for further tests.
Distinct colonies showing metallic green sheen on EMB were then subcultured on BCIG-SMAC (5-bromo-4chloro-3-indolyl-D-glucuronide-Sorbitol MacConkey) supplemented with CT (cefixime tellurite)38. E. coli O157:H7 monovalent antiserum was used for confirmation for confirmation of O157:H7.
Biofilm studies: Overnight pure cultures of 3 isolates of E. coli O157:H7 and a reference strain E. coli ATCC 35218 were used for biofilm development. Biofilm quantification was done using the crystal violet binding assay. The effect of the plant extracts on biofilm formation and biofilm dispersal was tested at 37°C for 48 and 24 hrs, respectively. The experiment was done in triplicates16.
Biofilm inhibition quantification: A modified crystal violet assay was employed to test the effect of plant extract on biofilm formation as earlier described39,40. The effect of extracts on biofilm formation was evaluated in 96-well polystyrene flat-bottom microtitre plates. A 50 μL of fresh bacterial suspension in modified tryptone soy broth (mTSB) at 108 dilution was aliquoted into each well of microplate and cultured in presence of 50 μL of the extract at 50 mg mL–1 concentration. Plates were incubated at 37°C for 48 hrs. After incubation, the supernatant was removed and each well was washed thoroughly with sterile distilled water to remove free-floating cells; thereafter plates were air-dried for 30 min, heat-fixed for a minute and the biofilm formed was stained during 15 min at room temperature with 0.1% aqueous solution of crystal violet. The excess stain was removed washing the plate three times with sterile distilled water. Finally, the dye bound to the cells was solubilized by adding 200 μL of 95% ethanol to each well and after 15 min of incubation at room temperature, absorbance was measured using a microplate reader at wavelengths of 432 and 630 nm. Tests were performed in triplicate.
Biofilm dispersion quantification: A modified crystal violet assay was employed to quantify the effect of plant crude extracts and fractions on biofilm formation as earlier described39,40. The effect of extracts on biofilm dispersion was evaluated in 96-well polystyrene flat-bottom microtitre plates. A 50 μL of fresh bacterial suspension in modified tryptone soy broth (mTSB) at 108 dilutions was aliquoted into each well of microplate and incubated at 37°C for 48 hrs. The plates were brought out and 50 μL of the 50 mg mL–1 concentration of the extracts was then added and incubated for another 24 hrs to determine the dispersion of already formed biofilm. After incubation, the supernatant was removed and each well was washed thoroughly with sterile distilled water to remove free-floating cells; thereafter plates were air-dried for 30 min, heat-fixed for a minute and the biofilm formed was stained during 15 min at room temperature with 0.1% aqueous solution of crystal violet. Following incubation, the excess of stain was removed washing the plate three times with sterile distilled water. Finally, the dye bound to the cells was solubilized by adding 200 μL of 95% ethanol to each well and after 15 min of incubation at room temperature, absorbance was measured using a microplate reader at a wavelength of 432 and 630 nm. Tests were performed in triplicate.
Statistical analysis: The experiment was replicated and every test was performed in triplicate. Data were analyzed using descriptive statistics and ANOVA at α 0.05.
RESULTS
Table 1 shows the biofilm formation inhibitory effects of all the tested extracts and crude while Table 2 shows the effect of how well the formed biofilms were dispersed.
Table 1 and 2 show that all isolates developed biofilms lies in a range of 0.08±0.07.
Biofilm development was inhibited by ethanolic (0.01±0.00) and crude extracts (0.05±0.01). The ethanolic and crude fractions had significant effects on biofilm formation inhibition followed by the ethyl acetate and hexane fractions while the chloroform fraction did not affect biofilm formation inhibition. The ethanolic fraction had the highest biofilm inhibition effect as seen in Table 1.
| Table 1: | Biofilm Inhibition of E. coli O157:H7 by extracts of ginger (Zingiber officinale) |
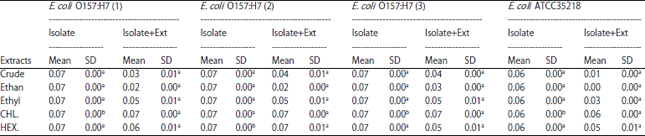 | |
SD: Standard deviation, Means in the same row not followed by the same lowercase letter are statistically significant (α<0.05) | |
| Table 2: | Biofilm dispersion of E. coli O157:H7 by extracts of ginger (Zingiber officinale) |
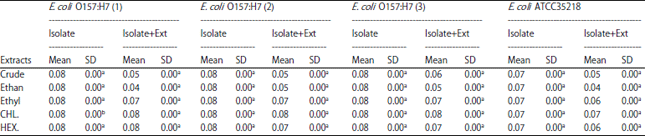 | |
SD: Standard deviation, Means in the same row not followed by the same lowercase letter are statistically significant (α<0.05) | |
In Table 2, the highest biofilm dispersion was exhibited with the ethanolic (0.04±0.05) and crude extract fractions (0.05±0.06). The chloroform fractions had little or no effect on biofilm formation and dispersal at 50 mg mL–1 concentration The ethanolic and crude fractions had a significant biofilm dispersion effect followed by the ethyl acetate and hexane fractions. The chloroform fraction did not affect biofilm dispersion. However, the ethanolic fraction had the highest biofilm dispersion effect as seen in Table 2.
The means by which microorganisms are inhibited by essential oils seem to involve different modes of action. Phenolic components present in essential oils have been known to possess antimicrobial activity and some are classified as Generally Recognized as Safe (GRAS) substances and therefore could be used to prevent post-harvest growth of native and contaminant bacteria. This study showed that ginger extract inhibited biofilm formation and also dispersed formed biofilm. Phenolic components of oils sensitize the phospholipid bilayer of the cell membrane, causing an increase of permeability and leakage of vital intracellular constitutes or impairment of bacterial enzyme systems. The phenolic component of ginger could have been responsible for the biofilm inhibition and dispersion reported in this study.
The ability of flavonoids to inhibit Quorum Sensing (QS) provides a clue for the biofilm inhibition and dispersion action recorded in this study as phytochemical studies showed that all the fractions of the extract have flavonoids4. An earlier study41 reported no complete inhibition for S. typhimurium or E. coli O157:H7 in the presence of ginger water extract. Another study28 further reported that two extracts (Fresh Ginger Extract (FGE) and Cooked Ginger Extract) impaired but did not prevent adhesion of all tested strains including E. coli O157:H7 when the extracts were used at a concentration of 10 and 15 mg DTSS/ mL and that generally, adhesion of all studied strains was reduced by increasing the concentration of FGE and CGE. The FGE had a strong effect as an anti-adhesive agent against S. Typhimurium strains and E. coli O157:H7 compared with the CGE. This may be due to the high content of gingerols in FGE. The study also reported that the minimum anti-biofilm concentration (MABC) of FGE and CGE were 20 and 25 mg DTSS/mL, respectively and that the FGE and CGE did not show complete inhibition of bacterial growth, although, it had considerable anti-adhesive and anti-biofilm effects.
Ginger extract showed the ability to inhibit Pseudomonas aeruginosa PA14 biofilm formation28, where biofilm development was reduced by 39-56% in a static biofilm assay using microtiter plates. The results obtained in this study demonstrated higher anti-biofilm potential of the ethanolic and crude extracts as compared with the ethyl acetate, hexane and chloroform extracts. This may be due to the high content of 6-, 8-, 10 gingerls as bioactive components in the ethanolic and crude extracts compared with others. In this respect, high level of 6-gingerol in the ethanolic and crude extracts has reduced biofilm formation and virulence by antagonistically binding to E. coli O157:H7 Quorum Sensing (QS) receptors42. Previously reported43 that a tested E. coli isolate exhibited high level of anti-spices resistance and that after exposure to a certain dose of spices, the isolate adapted biofilm mode of growth and with increasing concentration of spices, the thickness of the biofilm increased with increase in time and the extracellular biofilm matrix was found trapping the particles of spices used to induce the biofilm mode of growth. Heterogenous biofilm formation was noticed as all the cells were not found to produce extracellular matrix required for biofilm formation, some cells were embedded into the extracellular matrix while some cells were attached on the outer side of the matrix material. Although the spices mix contained NaCl and other compounds such as ginger and garlic, it was reported that the NaCl could have been responsible for the induction of biofilm mode of growth in the bacteria44-46 because salts generate high osmotic pressure which may damage the bacterial cell wall. Researcher47 described that the cell wall deformity and the state of cell wall deficiency encourage the biofilm formation in bacteria and protects it from toxic effect of antibacterial agents. Ginger and garlic are reported to have very strong antimicrobial activity48 and inhibit bacterial biofilms by blocking quorum sensors systems49. However, the composite spice mixes used for the study does not have the property to inhibit or kill bacteria because very small sub-inhibitory quantities of various herbs and spices were added to create the mix and they are used mainly as flavorings and coloring agents43.
Ginger extract was tested by Kim28 against P. aeruginosa PA14 biofilm formation and biofilm inhibition was demonstrated. A study on P. aeruginosa50 found out that the ethanolic extract of Z. officinale inhibited P. aeruginosa biofilm formation under both aerobic and anaerobic environments. A recent study with the phenolic compounds isolated from Z. officinale being QSI (quorum sensing inhibitors) was confirmed on P. aeruginosa MTCC 229751. That is important because quorum sensing is playing a significant role in food spoilage, biofilm formation, food-related pathogenesis and ginger is well used in foodstuff as a spice in dry or fresh form. Nikolic8 obtained similar results with this current study stating that the ethanolic extract of Zingiber officinale demonstrated moderate anti-biofilm activity and the influence was best on P. mirabilis were the BIC50 was at 19 mg mL–1. This study has implications for food safety and antibiotic resistance in biofilm-forming bacterial isolates.
Bacteria encountered in the food processing environment can be difficult to eliminate because of their adaptability under stress conditions and E. coli being one of such bacteria has been known to produce exopolysaccharides which can provide a physical barrier to protect the cell from environmental stress. Generally, biofilms are resistant to antibacterial agents. In food hygiene, biofilms are a very important potential hazard and a source of bacterial contamination, pre and post-production. Ginger (Zingiber officinale) showed potential use for dispersion of already formed biofilm by E. coli O157:H7 strains, hence, can be used in food processing, surfaces and industries to inhibit E. coli O157:H7 biofilm formation, increase dispersal and enhance food safety.
SIGNIFICANCE STATEMENT
This study discovered the biofilm inhibition and dispersion property of ginger (Zingiber officinale) that can be beneficial in food processing and the environment thus reducing antimicrobial resistance and enhance food safety. This study will help researchers to uncover the critical areas of the use of natural antimicrobials and anti-biofilm agents in food processing which many researchers were not able to explore. Thus, a new theory on natural anti-biofilm agents may be arrived at.
REFERENCES
- Donlan, R.M., 2002. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis., 8: 881-890.
CrossRefPubMedDirect Link - Park, M., J. Bae and D.S. Lee, 2008. Antibacterial activity of (10)-Gingerol and (12)-Gingerol isolated from Ginger Rhizome against periodontal bacteria. J. Phytother. Res., 22: 1446-1449.
CrossRef - Kociolek, M., 2009. Quorum-sensing inhibitors and biofilms. Antiinfect. Agents. J. Med. Chem., 8: 315-326.
CrossRefDirect Link - Carpentier, B. and O. Cerf, 1993. Biofilms and their consequences, with particular reference to hygiene in the food industry. J. Appl. Bacteriol., 75: 499-511.
CrossRefDirect Link - Kumar, C.G. and S.K. Anand, 1998. Significance of microbial biofilms in food industry: A review. Int. J. Food Microbiol., 42: 9-27.
CrossRefDirect Link - Rumbaugh, K.P. and I. Ahmad, 2014. Antibiofilm Agents. Springer, Berlin Heidelberg, Pages: 497.
CrossRefDirect Link - Nikolic, M., S. Vasic, J. Djurdjevic, O. Stefanovic and L. Comic, 2014. Antibacterial and anti-biofilm activity of ginger (Zingiber officinale (Roscoe)) ethanolic extract. Kragujevac J. Sci., 36: 129-136.
CrossRefDirect Link - Lim, J.Y., J.W. Yoon and C.J. Hovde, 2010. A brief overview of Escherichia coli O157:H7 and its plasmid O157. J. Microbiol. Biotechnol., 20: 5-14.
Direct Link - Melton-Celsa, A., K. Mohawk, L. Teel and A. OBrien, 2011. Pathogenesis of shiga-toxin producing Escherichia coli. In: Ricin and Shiga Toxins, Mantis, N., Springer, Berlin Heidelberg, pp: 67-103.
CrossRefDirect Link - Schuenzel, K.M. and M.A. Harrison, 2016. Microbial antagonists of foodborne pathogens on fresh, minimally processed vegetables. J. Food Prot., 65: 1909-1915.
CrossRefDirect Link - Frankel, G. and A.D. Phillips, 2008. Attaching effacing Escherichia coli and paradigms of Tir-triggered actin polymerization: getting off the pedestal. Cell Microbiol., 10: 549-556.
CrossRefDirect Link - Kalliomaki, M.A. and W.A. Walker, 2005. Physiologic and pathologic interactions of bacteria with gastrointestinal epithelium. Gastroenterol. Clin. North Am., 34: 383-399.
CrossRefPubMedDirect Link - Vogt, R.L. and L. Dippold, 2017. Escherichia coli O157:H7 outbreak associated with consumption of ground beef, June–July 2002. Public Health Rep., 120: 174-178.
CrossRefDirect Link - Larson, E.L., C. Gomez-Duarte, L.V. Lee, P. Della-Latta, D.J. Kain and B.H. Keswick, 2003. Microbial flora of hands of homemakers. Am. J. Infect. Control, 31: 72-79.
CrossRefDirect Link - Uhlich, G.A., P.H. Cooke and E.B. Solomon, 2006. Analyses of the red-dry-rough phenotype of an Escherichia coli O157:H7 strain and its role in biofilm formation and resistance to antibacterial agents. Appl. Environ. Microbiol., 72: 2564-2572.
CrossRefDirect Link - Garrett, T.R., M. Bhakoo and Z. Zhang, 2008. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci., 18: 1049-1056.
CrossRefDirect Link - Wojnicz, D., A.Z. Kucharska, A. Sokol-Letowska, M. Kicia and D. Tichaczek-Goska, 2012. Medicinal plants extracts affect virulence factors expression and biofilm formation by the uropathogenic Escherichia coli. Urol. Res., 40: 683-697.
CrossRefDirect Link - Cegelski, L., G.R. Marshall, G.R. Eldridge and S.J. Hultgren, 2008. The biology and future prospects of antivirulence therapies. Nat. Rev. Microbiol., 6: 17-27.
CrossRefDirect Link - OHara, C.M., C.D. Steward, J.L. Wright, F.C. Tenover and J.M. Miller, 1998. Isolation of Enterobacter intermedium from the gallbladder of a patient with cholecystitis. J. Clin. Microbiol., 36: 3055-3056.
Direct Link - Sang, S., J. Hong, H. Wu, J. Liu and C.S. Yang et al., 2009. Increased growth inhibitory effects on human cancer cells and anti-inflammatory potency of shogaols from Zingiber officinale relative to gingerols. J. Agric. Food Chem., 57: 10645-10650.
CrossRef - Shao, X., L. Lv, T. Parks, H. Wu, C.T. Ho and S. Sang, 2010. Quantitative analysis of ginger components in commercial products using liquid chromatography with electrochemical array detection. J. Agric. Food Chem., 58: 12608-12614.
CrossRefDirect Link - Sebiomo, A., A.D. Awofodu, A.O. Awosanya, F.E. Awotona and A.J. Ajayi, 2011. Comparative studies of antibacterial effect of some antibiotics and ginger (Zingibre officinale) on two pathogenic bacteria. J. Microbiol. Antimicrobials, 3: 18-22.
Direct Link - Betoni, J.E.C., R.P. Mantovani, L.N., Barbosa, L.C. Di Stasi and A. Fernandes Jr., 2006. Synergism between plant extract and antimicrobial drugs used on Staphylococcus aureus diseases. Mem. Inst. Oswaldo Cruz, 101: 387-390.
CrossRefDirect Link - Meliani, A., S. Nair and A. Bensoltane, 2014. Cyto-biochemical and antimicrobial investigations on essential oil of Zingiber officinale Roscoe. J. Essential Oil Bearing Plants, 17: 1120-1129.
CrossRefDirect Link - Maekawa, L.E., R.D. Rossoni, J.O. Barbosa, A.O.C. Jorge, J.C. Junqueira and M.C. Valera, 2015. Different extracts of zingiber officinale decrease enterococcus faecalis infection in galleria mellonella Braz. Dent. J. 26: 105-109.
CrossRefDirect Link - Kim, H.S. and H.D. Park, 2013. Ginger extract inhibits biofilm formation by Pseudomonas aeruginosa PA14. PLoS ONE, Vol. 8.
CrossRefDirect Link - Lee, K., J.H. Lee, S.I. Kim, M.H. Cho and J. Lee, 2014. Anti-biofilm, anti-hemolysis, and anti-virulence activities of black pepper, cananga, myrrh oils, and nerolidol against Staphylococcus aureus. Appl. Microbiol. Biotechnol., 98: 9447-9457.
CrossRefDirect Link - Agarwal, V., P. Lal and V. Pruthi, 2008. Prevention of Candida albicans biofilm by plant oils. Mycopathologia, 165: 13-19.
CrossRefPubMedDirect Link - Ali, B.H., G. Blunden, M.O. Tanira and A. Nemmar, 2008. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol., 46: 409-420.
CrossRefPubMedDirect Link - Skandamis, P., K. Koutsoumanis, K. Fasseas, and G.J.E. Nychas, 2001. Inhibition of oregano essential oil and EDTA on E. coli O157:H7. Italian J. Food Sci., 13: 55-65.
Direct Link - Karatan, E. and P. Watnick, 2009. Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiol. Mol. Biol. R, 73: 310-347.
CrossRefDirect Link - Miller, M.B. and B.L. Bassler, 2001. Quorum sensing in bacteria. Annu. Rev. Microbiol., 55: 165-199.
CrossRefDirect Link - Hegde, M., T.K. Wood and A. Jayaraman, 2009. The neuroendocrine hormone norepinephrine increases Pseudomonas aeruginosa PA14 virulence through the las quorum-sensing pathway. Appl. Microbiol. Biotechnol., 84: 763-776.
CrossRefDirect Link - Kong, K.F., C. Vuong and M. Otto, 2006. Staphylococcus quorum sensing in biofilm formation and infection. Int. J. Med. Microbiol., 296: 133-139.
CrossRefDirect Link - Xu, L., H. Li, C. Vuong, V. Vadyvaloo and J. Wang et al., 2006. Role of the luxS quorum-sensing system in biofilm formation and virulence of Staphylococcus epidermidis. Infect. Immun., 74: 488-496.
CrossRefDirect Link - Ryu, J.H. and L.R. Beuchat, 2005. Biofilm Formation by Escherichia coli O157: H7 on stainless steel: Effect of exopolysaccharide and curli production on its resistance to chlorine. Applied Environ. Microbiol., 71: 247-254.
CrossRefDirect Link - Adetunji, V.O. and G.O. Adegoke, 2008. Formation of biofilm by strains of Listeria monocytogenes isolated from soft cheese wara and its processing environment. Afr. J. Biotechnol., 7: 2893-2897.
Direct Link - Khiralla, G.M., 2015. Antibiofilm and anti-adhesive effects of ginger against some food-related pathogens. J. Food Res. Technol., 3: 87-96.
Direct Link - Indu, M.N., A.A.M. Hatha, C. Abirosh, U. Harsha and G. Vivekanandan, 2006. Antimicrobial activity of some of the South-Indian spices against serotypes of Eschrichia coli, Salmonella, Listeria monocytogenes and Aeromonas hydrophila. Braz. J. Microbiol., 37: 153-158.
CrossRefDirect Link - Kim, H.S., S.H. Lee, Y. Byun and H.D. Park, 2015. 6-Gingerol reduces Pseudomonas aeruginosa biofilm formation and virulence via quorum sensing inhibition. Sci. Rep., Vol. 5.
CrossRefDirect Link - Zulfiqar A.M., N.K. Muhammad, S. Anila, K. Fouzia and A. Mubashir et al., 2018. Ascorbic acid augments colony spreading by reducing biofilm formation of methicillin-resistant Staphylococcus aureus. Iran J. Basic Med. Sci., 21: 175-180.
CrossRefDirect Link - Lim, Y., M. Jana, T.T. Luong and C.Y. Lee, 2004. Control of glucose- and NaCl-Induced biofilm formation by rbf in Staphylococcus aureus. J. Basic Microbiol., 186: 722-729.
CrossRefDirect Link - Conlon, K.M., H. Humphreys and J.P. O'Gara, 2002. icaR encodes a transcriptional repressor involved in environmental regulation of ica operon expression and biofilm formation in Staphylococcus epidermidis. J. Bacteriol., 184: 4400-4408.
CrossRefPubMedDirect Link - Rachid, S., K. Ohlsen, U. Wallner, J. Hacker, M. Hecker and W. Ziebuhr, 2000. Alternative transcription factor ςB Is involved in regulation of biofilm expression in a Staphylococcus aureus mucosal isolate. J. Bacteriol., 182: 6824-6826.
CrossRefDirect Link - Mirani, Z.A. and N. Jamil, 2011. Effect of sub-lethal doses of vancomycin and oxacillin on biofilm formation by vancomycin intermediate resistant Staphylococcus aureus. J. Basic Microbiol., 51: 191-195.
CrossRefDirect Link - Shaista, J.K., F. Aneela, Z. Nasreen and E. Nusrat, 2009. Effect of different stabilizers on the antibacterial activity of “ginger garlic paste”. J. Appl. Sci. Environ. Manag., 13: 105-109.
CrossRefDirect Link - Musthafa, K.S., A.V. Ravi, A. Annapoorani, I.S.V. Packiavathy and S.K. Pandian, 2010. Evaluation of anti-quorum-sensing activity of edible plants and fruits through inhibition of the n-acyl-homoserine lactone system in Chromobacterium violaceum and Pseudomonas aeruginosa. Chemotherapy, 56: 333-339.
CrossRefPubMed - Yahya, M.F.Z.R., N.F.H.A. Saifuddin and U.M.A. Hamid, 2013. Zingiber officinale ethanolic extract inhibits formation of Pseudomonas aeruginosa biofilm. Int. J. Pharm. Biol. Sci., 3: 46-54.
Direct Link - Kumar, N.V., P.S. Murthy, J.R. Manjunatha and B.K. Bettadaiah, 2014. Synthesis and quorum sensing inhibitory activity of key phenolic compounds of ginger and their derivatives. Food Chem., 15: 451-457.
CrossRefDirect Link