
Gamal Mohamed Hamad
Department of Food Technology, Arid Land Cultivation Research Institute (ALCRI), City of Scientific Research and Technology Applications (SRTA-CITY), New Borg El-Arab City, 21934 Alexandria, Egypt
LiveDNA: 20.20321
Tarek Hosny Taha
Department of Environmental Biotechnology, Genetic Engineering and Biotechnology Research Institute (GEBRI), City of Scientific Research and Technology Applications (SRTA-CITY), New Borg El-Arab City, 21934 Alexandria, Egypt
Ali Mohammed Alshehri
Department of Biology, Faculty of Science, King Khalid University, 9004 Abha, Saudi Arabia
Elsayed Elsayed Hafez
Department of Plant Protection and Biomolecular Diagnosis, Arid Land Cultivation Research Institute (ALCRI), City of Scientific Research and Technology Applications (SRTA-CITY), New Borg El-Arab City, 21934 Alexandria, Egypt
Research Journal of Microbiology
Year: 2018 | Volume: 13 | Issue: 1 | Page No.: 28-36







ABSTRACT
Background and Objective: Production of glutathione through mutated yeast strains and the possible usage as antibacterial agent against some food borne bacteria. Also, the antioxidant activity of the glutathione considered as another value when added as food supplement and food preservative. Moreover development of a commercial medium mostly consists of secondary products for the high yeast propagation and high amount of the produced glutathione. The aim of this study was producing a mutated strains of Saccharomyces cerevisiae (S. cerevisiae) and Candida utilis (C. utilis); have high capability to produce high amount of glutathione. Materials and Methods: In this regard, two yeast strains; Saccharomyces cerevisiae and Candida utilis were examined for glutathione production. For increasing the production of the two yeast strains; chemical mutation using Ethyl methanesulfonate (EMS) was approached and the mutated strains were selected on medium contains the glutathione analogue. Comparison of means was performed with Duncan's multiple range test using Costat software. Results: Only two mutated strains showed high ability to produce glutathione with 49 folds more than wild types (MG40/S.C/4 and MG20/C.U/5). Moreover, the antioxidant capacity for the two mutated strains was 9 folds increased compared to wild types. The fermentation process was performed to analyze different parameters and it was observed that the medium should contains molasses as carbon source, yeast extract as a nitrogen source, KH2PO4 of mineral and cysteine for amino acid. Conclusion: Glutathione could be used as antioxidant and antibacterial against wide range of human pathogens bacteria in addition used as food additive, supplement and food preservation to control human pathogenic bacteria.
PDF Abstract XML References Citation
Received: August 16, 2017;
Accepted: October 23, 2017;
Published: February 23, 2019
How to cite this article
Gamal Mohamed Hamad, Tarek Hosny Taha, Ali Mohammed Alshehri and Elsayed Elsayed Hafez, 2018. Enhancement of the Glutathione Production by Mutated Yeast Strains and its Potential as Food Supplement and Preservative. Research Journal of Microbiology, 13: 28-36.
URL: https://scialert.net/abstract/?doi=jm.2018.28.36
URL: https://scialert.net/abstract/?doi=jm.2018.28.36
INTRODUCTION
Glutathione is a tripeptide compound consists of glutamic acid, cysteine and glycine and it is presented in most living organisms1,2. This compound plays an essential role in the metabolism of some toxic compounds and it has been used as a drug for hepatic insufficiency in humans, dogs and fish. In addition, for its importance as medicinal compound, glutathione was used also as an antiaging compound3. Glutathione peroxidase eliminates the active oxygen which resulted in both lipids and protein metabolism4,5,6. Zhang and Duan 7, Kipp et al.8 and Millman et al.9 reported that, the importance of the glutathione dived from its ability as antioxidative, immune inducer and detoxifying agents for some toxic compounds evolved in the cells. For that reasons, glutathione tablets and dried-yeast (producing glutathione) is widely used as food supplements in many different countries.
Glutathione (GSH) possesses life-sustaining functions as it is an important antioxidant synthesized by mammals. The deficiency of GSH has been reported to cause several diseases10,11 due to the oxidative stress12. GSH a nonprotein thiol compound is found in cells at the concentration of 1-10 mM. GSH is found to be the most important antioxidant, because of its antioxidizing capability is 100 times more than simple antioxidants. Therefore, there is a high demand and research on production of synthetic antioxidant13-16.
Glutathione (GSH) plays important roles in pulmonary diseases and inhaled GSH therapy has been used to treat cystic fibrosis (CF) patients in clinical trials. The results in this report revealed that GSH altered the sensitivity of Pseudomonas aeruginosa (P. aeruginosa) to different antibiotics through pathways unrelated to the oxidative stress as generally perceived. In addition, GSH and its oxidized form inhibited the growth of P. aeruginosa17. The GSH show high antibacterial activity agents Pseudomonas aeruginosa; Enterobacter; Staphylococcus aureus; Micrococcus luteus; Klebsiella pneumoniae and Escherichia coli18.
The process of glutathione extraction from yeast cells is one of the inexpensive methods. In Japan, Candida utilis was used for production of glutathione since 196019. Several studies were approached using both of S. cerevisiae and C. utilis for glutathione production and the optimization conditions which used to increase the glutathione productivity by the addition of amino acids, especially cysteine20. Different studies used glucose and ethanol21,22 and optimal controls of oxygen and pH values12 resulted in high productivity. In addition methanol was used as sole carbon source for glutathione production by Pichia pastoris and Hansenula polymorpha23-26.
Saccharomyces cerevisiae is easy eukaryotic organism to manipulate genetically and it produces a high amount of glutathione but there was little information about glutathione synthesis and regulation27,28. It was reported that S. cerevisiae could be inhibited by methylglyoxal28. Angelov et al.28 reported that the mutant cells deficient in glyoxalase I are more sensitive to methylglyoxal than the wild type cells and the lack of glutathione resulted in deficiency of the glyoxalase system since the increase in yeast increase sensitivity to methylglyoxal. The main aims of this study to produce glutathione from two yeast strains; Saccharomyces cerevisiae ATCC 9763 and Candida utilis ATCC 9950 and improved the strains glutathione production by mutation.
Ethyl methanesulfonate (EMS) has the formula C3H8SO3 and considered as mutagenic and possibly carcinogenic compound. Its mutagenic principle is depending on nucleotide substitution especially guanine alkylation and so it only cause a point mutation. The mutated gene could increase its production rate up to 5×10–2 without harm and undesired effects on the gene. It is mainly depending on place of thymine instead of cytosine that opposite O-6-ethylguanine (formed by reaction with the ethyl group of EMS) during DNA replication. The new formed nucleotide pair could be A:T instead of G:C. The new formed mutation could affect the behavior and the production rate of the mutated gene29,30. The aim of this study was producing a muated starians of Saccharomyces cerevisiae and Candida utilis; have high capability to produce high amount of glutathione. Tested the obtained glutathione as antioxidant and antibacterial against a wide range of human pathogenic bacteria as well.
MATERIALS AND METHODS
Chemicals and solutions: All chemicals and solutions used in this study are HPLC grade and purchased from Sigma-Aldresh Company (Germany).
Yeast strains and cultivation conditions: Yeast strains Saccharomyces cerevisiae DSM 70487 was obtained from The Department of Microbiology, Faculty of Agriculture, Ain Shams University, Cairo, Egypt, while Candida utilis EMCC 41 was obtained from Cairo MERCEN, Faculty of Agriculture, Ain Shams University, Cairo, Egypt. S. cerevisiae and C. utilis were obtained as lypholized materials, then dissolved in 10 mL distilled H2O and 10 μL of each dilution was seprated on solid Yeast Peptone Dextrose (YPD) medium free antibiotics. One colony from each strain was transferred and grown on broth YPD medium, 3 mL under aerobic conditions with shaking at 30°C (10 g of yeast extract, 5 g of peptone and 20 g of glucose L–1 18. S. cerevisiae and C. utilis were grown under aerobic conditions on rotary shaker (225 rpm) in YPD media (10 g of yeast extract, 5 g of peptone and 20 g of glucose L–1) at temperature 30°C.31
Induction of mutation and isolation of methylglyoxal resistant mutants: Mutations were induced by treatment of yeasts cells with ethyl methanesulfonate (EMS) with 2% concentration (20 mg mL–1) according to Shiomi et al.18 In summarized; cells were precultured in 10 mL of YPD medium for 3 days. The precultured cells were inoculated at a concentration of 0.1 (OD610) in 50 mL of YPD medium and cultured at 30°C with shaking at 120 rpm. After 1 day, the cells were suspended into a 2 mL microtube and washed 3 times with 10 mL of 0.1 M citric buffer (pH 5.5) and then cells were collected by centrifugation at 5000 rpm for 2 min, resuspended cells in 10 mL of buffer containing 2% ethyl methanesulfonate (EMS) and 5% glucose. The cultures were shaken and then incubated at 30°C for 1 h. The cells were washed 3 times with 1.0 mL of 0.1 M citric buffer (pH 5.5). The treated cells were spread on agar plates of YPD medium. Later, all colonies grown on the plates were collected and diluted and 1000 cells were spread on each agar plate of YPD medium containing 0.20 M methylglyoxal analogue. The plate was incubated at 30°C for 5 days and then colonies were isolated as methylglyoxal resistant mutants18.
Preparation of cells for glutathione production: The two yeast strains were prepared for the glutathione production18. Yeast strains were precultured in 20 mL YPD medium at 30°C for 3 days and washed with 20 mL of water and then were suspended in 1 mL water. The precultured cells were inoculated at optical densities (OD) of 0.1 at 610 nm into 100 mL of medium in a 500 mL flask and the cells were cultured for 3 days at 30°C with shaking at 120 rpm.
Glutathione extraction: Glutathione was extracted from the yeast cells according to Fan et al.31. The procedure was summarized in; after incubation cells were collected from centrifugation at 6400 rpm for 10 min and washed twice with distilled water and resuspended in 0.5 mL of distilled water. The cell suspension was incubated in a boiling water bath for 5 min and centrifugation at 6400 rpm for 10 min. The glutathione contents in the supernatant were determined by colorimetric method using 5, 5-Dithiobis (2-nitrobenzoic acid, DTNB).
Determination of glutathione in yeast culture filtrate: The glutathione was determined using colorimetric methods using glutathione reduced kit (Glutathione Reduced colorimetric kit, Biodiagnostic) by measuring the absorbance of reaction solutions at 405 nm using a spectrophotometer (Konica Minolta, Europ). One mililiter from sample was added to 0.5 mL distilled water and mixed with 0.5 mL the kit reagent 1. The soultion was mixed well and allowed to stand for 5 min and then centrifuged at 3000 rpm for 15 min. A 0.5 mL of supernatant was taken and mixed with 1 mL reagent 2 followed by addition of 100 μL reagent 3 and mixed well. After 10 min, the OD was measured at 405 nm against the blank (0.5 mL of mixture, 0.5 mL distilled water with 0.5 mL reagent 1 was added to 1 mL reagent 2 and 100 μL reagent 3). The glutathione concentration was calculated32 as = sample OD×66.6 = mg dL–1.
Fermentation optimization for glutathione production: For glutathione production enhancement both the yeast strains were grown in 500 mL flask containing 100 mL of yeast medium (YM) (1.0% glucose, 0.5% peptone, 0.3% yeast extract and 0.3% malt extract, at pH 6.0 for 24 h at 3°C)21. Culture cells were inoculated into 1 L flasks, each containing 200 mL of the same medium and then incubated at 3°C for 72 h with agitation at 100 rpm. After incubation, the culture was centrifuged at 7,000 rpm for 15 min, the supernatant removed and the yeast cells washed with distilled water 3 times. The harvested yeast cells analyzed for glutathione concentration and dry cell weight. Glutathione production under optimal culture conditions in YM was undertaken to investigate the influence of precursor amino acids (0.05% salt, (0.05%) carbon and nitrogen sources (1%) on glutathione production.
Antibacterial activity
Microorganisms and culture conditions: Pathogenic bacteria strains used were; four Gram-positive strains; Bacillus cereus EMCC 1006, Staphylococcus aureus EMCC 1351, Streptococcus pyogenes EMCC 1772 and Streptococcus mutans EMCC 1815 and three Gram-negative strains; Salmonella spp., Escherichia coli ATCC 25922., Klebsiella pneumoniae EMCC 1637 and Pseudomonas aeruginosa EMCC1256. All strains were obtained from Microbiological Resources Centres (MERCIN), Faculty of Agriculture, Ain Shams University, Cairo, Egypt. The strains were maintained in 60% glycerol/ LB culture at -80°C.
Effect of glutathione and minimum inhibitory concentration (MIC) determination: The antimicrobial activity of glutathione by agar well diffusion assay33 against pathogenic bacteria. The bacterial strains were grown in nutrient broth at 37°C. Briefly, 100 μL of overnight culture of each pathogens strain (106 CFU mL–1) were aseptically spread over LB agar plates. About 100 μL of sterile glutathione was transferred to each agar well. The plates were then incubated at 37°C for 18 h and the formed clear zones (if found) were measured and recorded. A set of 6 concentration of reconstituted plant water extracts (0.5, 0.3, 0.1, 0.05 and 0.025 mg mL–1), were examined to determine the Minimum Inhibitory Concentration (MIC) of each against a specific pathogenic strain.2 The zone of inhibition was calculated by measuring the diameter of the inhibition zone around the well (mm), including the well diameter. The readings were taken in three different fixed directions in all triplicates and the average values were tabulated.
Total antioxidant determination of the obtained glutathione: Total Antioxidant Capacity (TAC) level was determined by using Biodiagnostic kit according to the manufacture process. The residual H2O2 was determined calorimetrically by an enzymatic reaction which involved the conversion of 3, 5-Dichloro-2-hydroxybenzensulphate to a colored product which can be measured at OD505 nm34.
Statistical analysis: Comparison of means was performed with Duncan's multiple range test (Duncan, 1955)35 at p<0.05 using the statistical analysis software 'CoStat 6.4' (CoStat, 2005)36.
RESULTS AND DISCUSSION
The production levels of glutathione are quantitatively determined and the results indicated that the original strain Saccharomyces cerevisiae (ATCC 9763) produced (16.5623 mg L–1) whenever the Candida utilis (ATCC 9950) gave 14.4325 mg L–1. The results revealed that S. cerevisiae is more productive than the C. utilis for glutathione by 12%. The highest amount of glutathione obtained by Saccharomyces cerevisiae ATCC 7754 was 25.1 mg37. Whenever, about 90 mg L–1 of glutathione was obtained by Saccharomyces cerevisiae FF-8 when cultivated on YM21. The previous studies and the obtained results revealed that the amount of glutathione vary from yeast strain to another.
After exposing the Saccharomyces cerevisiae strain to EMS, the mutated cells were cultivated on YPD medium containing glutathione analogue (methylglyoxal), it was observed that eighteen resistant mutants were obtained from 2% EMS treatment (Table 1). Moreover, the survival percentage was 55.8%. On the other hand, it was obvious that the increasing in methylglyoxal concentrations lead to decrease in the survival percentage and it was observed that the survival percentages; 0.39, 0.26 and 0.13% were obtained when the cells treated by concentrations of 20, 40 and 60 μg mL–1, respectively.
Results obtained by the mutated Candida utilis were slightly differences on which obtained by S. cerevisiae. The survival percentage was 55.2% for treatments by 2% EMS. The different treatments showed different survival as shown in (Table 1). Results presented in this study revealed that the concentration of 20 μg mL–1 is the best for the both examined fungal isolates. These results are similar to what obtained by previous studies38,18. The same results were obtained by Ohtake et al.39 when a mutagenesis was performed on Saccharomyces cerevisiae YNN 27 using EMS. They reported that 12 mutants were obtained and they classified these mutants into two classes.
Glutathione production by the mutant Saccharomyces cerevisiae: The production of glutathione by the mutant strain Saccharomyces cerevisiae were varied according to the concentration of the analogue concentration. The production of the glutathione was ranged from 0.01134-0.08048 g L–1 with EMS methylglyoxal (20 μg mL–1). Wherever, the production ranged from 0.02320-0.58330 g L–1 with methylglyoxal (40 μg mL–1) and it was 0.03308-0.05345 g L–1 with methylglyoxal (60 μg mL–1) (Table 2). Here in this study, it was obvious that the glutathione production by the mutant strain (MG40/S.C/4) was more 49 times than that obtained by the original strain.
| Table 1: | Survival percentages of methylglyoxal resistant mutants of Saccharomyces cerevisiae and Candida utilis |
 | |
| Table 2: | Production of glutathione (mg L–1) by EMS mutated S. cerevisiae treated with different concentrations of glutathione analogue (methylglyoxal) |
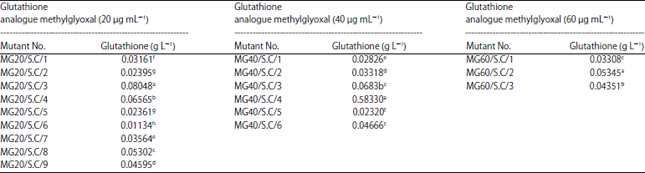 | |
Means with the same letter are not significantly different according to Duncan’s multiple range test least significant difference (LSD) 0.01 = 1.37 in glutathione analogue methylglyoxal (20 μg mL–1), LSD 0.01 = 1.4737 in glutathione analogue methylglyoxal (40 μg mL–1) and LSD 0.01 = 1.7937 in glutathione analogue methylglyoxal (60 μg mL–1). Each value represents the mean of 3 replicates | |
| Table 3: | Production of glutathione (mg L–1) by EMS mutated Candida utilis treated with different concentration of glutathione analogue (methylglyoxal) |
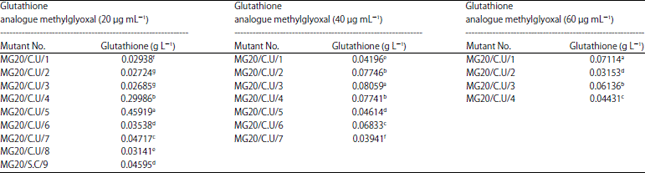 | |
Means with the same letter are not significantly different according to Duncan’s p<0.05 multiple range test least significant difference (LSD) 0.01 = 1.0629 in glutathione analogue methylglyoxal (20 μg mL–1), LSD 0.01 = LSD 0.01 = 1.2552 in glutathione analogue methylglyoxal (40 μg mL–1) and LSD 0.01 = 1.1343 in glutathione analogue methylglyoxal (60 μg mL–1), Each value represents the mean of 3 replicates | |
According to Angelov et al.28; they exposed the S. cerevisiae strain to the mutagenic EMS and they observed that the mutant strains were completely different on the wild type based on their glutathione production. The same observation was obtained by Nishiuchi et al.40.
Glutathione production capacity in Candida utilis: The results that presented in (Table 3), revealed that there are wide variations in glutathione production. The mutants Candida utilis that grew at 20 μg mL–1 of methylglyoxal gave production ranged between 0.02685 and 0.459.19 g L–1. Wherever, production ranged from 0.03941-0.08059 g L–1 were obtained by the mutants of (methylglyoxal 40 μg mL–1), also, the production was fluctuated from 0.03153-0.07114 g L–1 with methylglyoxal 60 μg mL–1 grown mutants. Here in this study, it was obvious that the glutathione production by the mutant strain (MG20/C.U/5) was more 31 times that obtained by the original strain. Shomai et al.18, reported that the mutated Pichia strains gave high glutathione production more than the wild types. These results were confirmed by what postulated by Magherini et al.40, that yeast can live on any carbon source either fermentable or non-fermentable. Beshay and Moreira41 reported that carbohydrate, nutrient, vitamins that are presented in molass are very essential for yeast performance.
Optimization conditions for high production of glutathione by the mutant strains: Different carbon sources were used to improve the fungal glutathione production and the results presented in (Table 4) revealed that molasses is the best carbon source. On contrary; Li et al.42 revealed that glucose is the best carbon source for glutathione production whenever, lactose reduce the glutathione production by 50% when compared with that obtained by glucose. For the nitrogen source, yeast extract showed the highest yield and this results was confirmed with what reported by Shin et al.43. The KH2PO4 is the mineral salt which gave high glutathione production when compared with other salts. The same observation was obtained by Cha et al.21 that KH2PO4 is best mineral salt with concentration 0.06% for high production of glutathione by S. cerevisiae. Moreover, it has been observed that, the presence of amino acids in the culture medium could increase the glutathione production compared with the absence of amino acids (Table 4). Li et al.42 reported that the addition of amino acids into the cultivation medium for the yeast increase the glutathione production by 1.4 fold. The same results obtained by Anschau et al.37 and they postulated that molass is the best carbon source for glutathione production when used as carbon source of the medium of Saccharomyces cerevisiae. In addition, the biomass of the yeast was increased when molass was used as carbon source when Anschau and his colleges added molasses and glycerol in the medium for increasing the glutathione production37.
But glucose is the best carbon source for glutathione production by the yeast strain Hansenula polymorpha DL-144. They reported that the production was increased from 900 up to 2300 mg L–1 concerning the importance of the salts in the medium, Ito et al.45 revealed that yeast intact cells could affect the gene expression when alkaline cations used in the cultivation medium. As well as they found that amino acids are essential for growth and glutathione production when both are supplied in the cultivation medium for yeast. The same observation was also reported by Beshay and Moreira41. In addition; it well known that the glutathione production controlled by enzyme pathway and this pathway affected by the substrate constitutions37. Anschau et al.37 found that the addition of amino acids into the yeast medium increased the production of glutathione from 236.1 mg L–1 after 96 h. Cha et al.21 reported that the high glutathione production was obtained when used the glucose 3.0% (w/v), yeast extract 3.0%, KH2PO4 0.06% and L-cysteine 0.06% in basal YM. Also, the addition of amino acids especially cysteine into the yeast medium increased the glutathione production46. Yamada et al.47, reported that Candida tropicalis pK 233 gave high amount of glutathione when they added ethanol as a sole carbon source in the cultivation medium. Abegg et al.48 revealed that Candida albicans when exposed to oxidative stress showed more production of glutathione.
Antibacterial activity and minimum inhibitory concentration (MIC): Antibacterial activity of glutathione was studied individually against tested pathogenic bacteria was exhibited in (Table 5), expressed by inhibition zone diameters and MICs (Minimum Inhibition Concentration).
| Table 4: | Effect of various medium constituents on both of glutathione production and cell dry weight of Saccharomyces cerevisiae (MG40/S.C/4) |
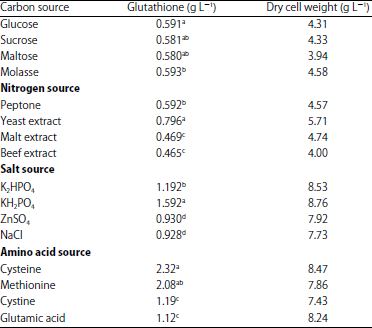 | |
Means with the same letter are not significantly different according to Duncan’s p<0.05 multiple range test (LSD 0.01 = 1.06, LSD 0.01 = 1.25, LSD 0.01 = 1.134). Each value represents the mean of 3 replicates | |
| Table 5: | Inhibition zones of glutathione (GSH) and minimum inhibitory concentration (MIC) determination |
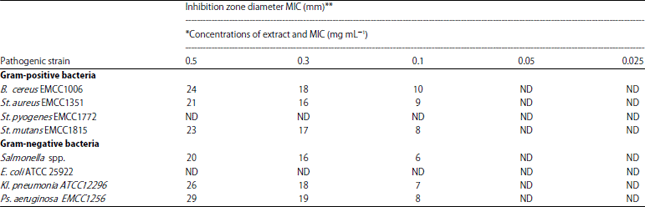 | |
| MIC: Minimum inhibition concentration. *Concentrations of extract and MIC are in mg mL–1. **Diameter included 5 mm well diameter. ND: Not detected | |
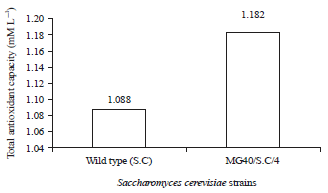 | |
| Fig. 1: | Total antioxidant capacity of glutathion produced by mutant Saccharomyces cerevisiae compared with wild type |
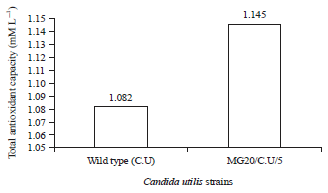 | |
| Fig. 2: | Total antioxidant capacity of glutathion produced by mutant Candida utilis compared with wild type |
Results showed good antibacterial activity against, Gram-positive strains, Bacillus cereus EMCC 1006, Staphylococcus aureus EMCC 1351, Streptococcus mutans EMCC 1815 and Bacillus cereus EMCC 1006 with MIC 0.05 mg mL–1, but glutathione did not show antibacterial effect on Streptococcus pyogenes EMCC 1772 and also the Gram-negative strains, Salmonella spp, Klebsiella pneumonia ATCC 12296 and Pseudomonas aeruginosa EMCC 1256 showed good antibacterial activity with MIC 0.05 mg mL–1 but glutathione did not show antibacterial effect on Escherichia coli ATCC 25922. These results are consistent with previous reports regarding "Gram-positive" and "Gram-negative" bacteria, which reported that the later are more resistant17.
Total antioxidant capacity: The total antioxidant capacity for the glutathione produced by Saccharomyces cerevisiae (Wild type and MG40/S.C/4 mutant strain) was listed in Fig. 1. However, the antioxidant obtained by Candida utilis (Wild type and MG20/S.U/5 mutant strain) it was lower than S. cerevisiae as shown in Fig. 2. According to Kerksick and Willough49 glutathione is considered strong antioxidant due to its ability to minimize oxidative stress and the lipid peroxidation of cellular membranes. The same opinion was mentiond by Priscilla and Heather 50 who reported that usage of the antioxidant substances such as glutathione will help in improvement of human health.
CONCLUSION
In this study, our wild type yeasts showed ability to produce glutathione with considerable amount not exceeded more than 17 mg L–1. The glutathione produced by yeast exposed to the mutagenesis and EMS up to 49 folds. The production of glutathione by the two examined yeast strains could be maximized if suitable fermentation medium of like molasses were approached. Glutathione showed no toxicity on the human cell and it has high antioxidant and antibacterial activity. Due to all the previous characters glutathione could be used as food additive and supplement to control human pathogenic bacteria.
SIGNIFICANCE STATEMENT
The study discovers that chemical mutation could affect the glutathione production by mutated yeast strains in compared with the wild ones. This study will help many researchers to do further studies in the importance of the glutathione as antibacterial and economic production of glutathione as food support.
REFERENCES
- Li, Y., G. Wei and J. Chen, 2004. Glutathione: A review on biotechnological production. Applied Microbiol. Biotechnol., 66: 233-242.
CrossRefDirect Link - Kern, J.K., A.G. David, B.A. James, R.G. Carolyn, A. Tapan and R.G. Mark, 2011. A clinical trial of glutathione supplementation in autism spectrum disorders. Med. Sci. Monit., 17: CR677-CR682.
CrossRefDirect Link - Weschawalit, S., S. Thongthip, P. Phutrakool and P. Asawanonda, 2017. Glutathione and its antiaging and antimelanogenic effects. Clin. Cosmet. Investig. Dermatol., 10: 147-153.
CrossRefDirect Link - Rahman, K., 2007. Studies on free radicals, antioxidants and co-factors. Clin. Interv. Aging, 2: 219-236.
Direct Link - Valko, M., M. Izakovic, M. Mazur, C.J. Rhodes and J. Telser, 2004. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem., 266: 37-56.
CrossRefDirect Link - Wu, G., Y.Z. Fang, S. Yang, J.R. Lupton and N.D. Turner, 2004. Glutathione metabolism and its implications for health. J. Nutr., 134: 489-492.
PubMedDirect Link - Zhang, Y. and K. Duan, 2009. Glutathione exhibits antibacterial activity and increases tetracycline efficacy against Pseudomonas aeruginosa. Sci. China Ser. C: Life Sci., 52: 501-505.
CrossRefDirect Link - Kipp, A., A. Banning and R. Brigelius-Flohe, 2007. Activation of the glutathione peroxidase 2 (GPx2) promoter by β-catenin. Biol. Chem., 388: 1027-1033.
CrossRefDirect Link - Millman, A.C., M. Salman and Y.K. Dayaram, 2008. Natural killer cells, glutathione, cytokines and innate immunity against Mycobacterium tuberculosis. J. Interferon Cytokine Res., 28: 153-165.
CrossRefDirect Link - Ballatori, N., S.M. Krance, S. Notenboom, S. Shi, K. Tieu and C.L. Hammond, 2009. Glutathione dysregulation and the etiology and progression of human diseases. Biol. Chem., 390: 191-214.
CrossRefPubMedDirect Link - Lu, S.C., 2009. Regulation of glutathione synthesis. Mol. Aspects Med., 30: 42-59.
CrossRefPubMedDirect Link - Valencia, E., A. Marin and G. Hardy, 2001. Glutathione-nutritional and pharmacological viewpoints: Part III. Nutrition, 17: 696-697.
CrossRefPubMedDirect Link - Xiong, Z.Q., M.J. Guo, Y.X. Guo, J. Chu, Y.P. Zhuang and S.L. Zhang, 2009. Efficient extraction of intracellular reduced glutathione from fermentation broth of Saccharomyces cerevisiae by ethanol. Bioresour. Technol., 100: 1011-1014.
CrossRefDirect Link - Das, A.J., S. Chalil, P. Nigam, P. Magee, O. Janneh and R. Owusu-Apenten, 2011. Glutathione transferase-P1-1 binding with naturally occurring ligands: assessment by docking simulations. J. Biophys. Chem., 2: 401-407.
CrossRefDirect Link - Gostimskaya, I. and C.M. Grant, 2016. Yeast mitochondrial glutathione is an essential antioxidant with mitochondrial thioredoxin providing a back-up system. Free Radical Biol. Med., 94: 55-65.
CrossRefDirect Link - Farid, A., A.H. Shah, M. Ayaz, A. Amin, M. Yaseen, H. Ullah and F. Haq, 2012. Comparative study of biological activity of glutathione, sodium tungstate and glutathione-tungstate mixture. Afr. J. Biotechnol., 11: 10431-10437.
CrossRefDirect Link - Shiomi, N., E. Harada and R. Nabeshima, 2012. Glutathione production by Yarrowia lipolytica showing both methyglyoxal resistance and a high activity of glutathione synthetase. Adv. Microbiol., 2: 171-180.
CrossRefDirect Link - Alfafara, C.G., K. Miura, H. Shimizu, S. Shioya and K.I. Suga, 1992. Cysteine addition strategy for maximum glutathione production in fed-batch culture of Saccharomyces cerevisiae. Applied Microbiol. Biotechnol., 37: 141-146.
CrossRefDirect Link - Schmacht, M., E. Lorenz, U. Stahl and M. Senz, 2017. Medium optimization based on yeast's elemental composition for glutathione production in Saccharomyces cerevisiae. J. Biosci Bioeng., 123: 555-561.
CrossRefDirect Link - Nie, W., G. Wei, G. Du, Y. Li and J. Chen, 2005. Enhanced intracellular glutathione synthesis and excretion capability of Candida utilis by using a low pH-stress strategy. Lett. Applied Microbiol., 40: 378-384.
CrossRefDirect Link - Fickers, P., P.H. Benetti, Y. Wache, A. Marty, S. Mauersberger, M.S. Smit and J.M. Nicaud, 2005. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica and its potential applications. FEMS Yeast Res., 5: 527-543.
CrossRefDirect Link - Shiomi, N., Y. Inoue, K. Tomioka and T. Yasuda, 2003. Immobilization of Candida cells showing mycelium-like shapes in porous polyvinyl formal resin and their applications. J. Chem. Eng. Jap., 36: 161-165.
CrossRefDirect Link - Gellissen, G., G. Kunze, C. Gaillardin, J.M. Cregg, E. Berardi, M. Veenhuis and I. van der Klei, 2005. New yeast expression platforms based on methylotrophic Hansenula polymorpha and Pichia pastoris and on dimorphic Arxula adeninivorans and Yarrowia lipolytica-a comparison. FEMS Yeast Res., 5: 1079-1096.
CrossRefDirect Link - Schmacht, M., E. Lorenz and M. Senz, 2017. Microbial production of glutathione. World J. Microbiol. Biotechnol., Vol. 33.
CrossRefDirect Link - Angelov, A., T. Hristozova and Z. Roshkova, 2000. Isolation of glutathione biosynthesis-deficient mutants of Saccharomyces cerevisiae and their use in baker's yeast production. Eur. Food Res. Technol., 211: 429-432.
CrossRefDirect Link - Arisha, M.H., S.N.M. Shah, Z.H. Gong, H. Jing, C. Li and H.X. Zhang, 2015. Ethyl methane sulfonate induced mutations in M2 generation and physiological variations in M1 generation of peppers (Capsicum annuum L.). Front. Plant Sci., Vol. 6.
CrossRefDirect Link - Caldwell, D.G., N. Mccallum, P. Shaw, G.J. Muehlbauer, D.F. Marshall and R. Waugh, 2004. A structured mutant population for forward and reverse genetics in Barley (Hordeum vulgare L.). Plant J., 40: 143-150.
CrossRefDirect Link - Fan, X., X. He, X. Guo, N. Qu, C. Wang and B. Zhang, 2004. Increasing glutathione formation by functional expression of the γ-glutamylcysteine synthetase gene in Saccharomyces cerevisiae. Biotechnol. Lett., 26: 415-417.
CrossRefDirect Link - Hara, K.Y., N. Aoki, J. Kobayashi, K. Kiriyama, K. Nishida, M. Araki and A. Kondo, 2015. Improvement of oxidized glutathione fermentation by thiol redox metabolism engineering in Saccharomyces cerevisiae. Applied Microbiol. Biotechnol., 99: 9771-9778.
CrossRefDirect Link - Kadaikunnan, S., T. Rejiniemon and J.M. Khaled, 2015. In-vitro antibacterial, antifungal, antioxidant and functional properties of Bacillus amyloliquefaciens. Ann. Clin. Microbiol. Antimicrob., Vol. 14.
CrossRefDirect Link - Koracevic, D., G. Koracevic, V. Djordjevic, S. Andrejevic and V. Cosic, 2001. Method for the measurement of antioxidant activity in human fluids. J. Clin. Pathol., 54: 356-361.
CrossRefPubMedDirect Link - Anschau, A., L.O. dos Santos and R.M. Alegre, 2013. A cost effective fermentative production of glutathione by Saccharomyces cerevisiae with cane molasses and glycerol. Braz. Arch. Biol. Technol., 56: 849-857.
CrossRefDirect Link - Nishiuchi, H., Y. Tabira and K. Yamagishi, 2012. A combination of flow cytometry and traditional screening using chemicals to isolate high glutathione-producing yeast mutants. Biosci. Biotechnol. Biochem., 76: 1085-1090.
CrossRefDirect Link - Ohtake, Y., A. Satou and S. Yabuuchi, 1990. Isolation and characterization of glutathione biosynthesis-deficient mutants in Saccharomyces cerevisiae. Agric. Biol. Chem., 54: 3145-3150.
CrossRefDirect Link - Magherini, F., A. Carpentieri, A. Amoresano, T. Gamberi, C. de Filippo and L. Rizzetto, 2009. Different carbon sources affect lifespan and protein redox state during Saccharomyces cerevisiae chronological ageing. Cell. Mol. Life Sci., 66: 933-947.
CrossRefDirect Link - Beshay, U. and A. Moreira, 2005. Production of alkaline protease with Teredinobacter turnirae in controlled fed-batch fermentation. Biotechnol. Lett., 27: 1457-1460.
CrossRefDirect Link - Li, Y., J. Chen, Y. Mao, S. Lun and Y.M. Koo, 1999. Fermentation conditions for production of glutathione by recombinant Escherichia coli. Wei Sheng Bao Xue Bao, 39: 355-361, (In Chinese).
PubMedDirect Link - Ubiyvovk, V.M., V.M. Ananin, A.Y. Malyshev, H.A. Kang and A.A. Sibirny, 2011. Optimization of glutathione production in batch and fed-batch cultures by the wild-type and recombinant strains of the methylotrophic yeast Hansenula polymorpha DL-1. BMC Biotechnol., Vol. 11.
CrossRefDirect Link - Ito, H., Y. Fukuda and A. Kimura, 1983. Transformation of intact yeast cells treated with alkali cations. J. Bacteriol., 153: 163-168.
PubMedDirect Link - Wen, S., T. Zhang and T. Tan, 2005. Optimization of the amino acid composition in glutathione fermentation. Process Biochem., 40: 3474-3479.
CrossRefDirect Link - Yamada, Y., Y. Tanil and T. Kamihara, 1984. Production of extracellular glutathione by Candida tropicalis Pk 233. J. Gen. Microbiol., 130: 3275-3278.
CrossRefDirect Link - Abegg, M.A., P.V.G. Alabarse, A.K. Schuller and M.S. Benfato, 2012. Glutathione levels in and total antioxidant capacity of Candida sp. cells exposed to oxidative stress caused by hydrogen peroxide. Rev. Soc. Bras. Med. Trop., 45: 620-626.
CrossRefDirect Link - Kerksick, C. and D. Willoughby, 2005. The antioxidant role of glutathione and N-acetyl-cysteine supplements and exercise-induced oxidative stress. J. Int. Soc. Sports Nutr., Vol. 2.
CrossRefDirect Link - Priscilla, C.M. and S.T. Heather, 2000. Antioxidants: What role do they play in physical activity and health? Am. J. Clin. Nutr., 72: 637s-646s.
Direct Link