
P. Gupta
Department of Biotechnology, National Institute of Technology, Raipur-492 010, India
L.S.B. Upadhyay
Department of Biotechnology, National Institute of Technology, Raipur-492 010, India
R. Shrivastava
Department of Biotechnology, National Institute of Technology, Raipur-492 010, India
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 281-288







ABSTRACT
Lipase-catalyzed transesterification of vegetable oils is an important reaction that produces fatty acid alkyl esters which are valuable intermediates in oleo chemistry and are excellent substitutes to diesel fuel. In present work lipase producing bacteria were isolated from oil spilled soil samples collected from different areas of Raipur, India by serial dilution method. Lipase activity of extracellular lipase was determined by titrimetric method. The sodium alginate entrapment was carried out to immobilize lipase according to the standard method. Out of 15 bacterial isolates (LPB1-LPB15), LPB1 exhibited the maximum extracellular lipase activity on lipase assay medium. Thus, it was selected for further study. Olive oil was found to be the best substrate for lipase production (0.0070 μg/mL /min) among the substrates tested. This isolate exhibited further increase in activity with value of 0.0099 μg/mL/min using olive oil as substrate in production medium supplemented with lecithin as emulsifier at pH 7.2 after 3 days of incubation at 30°C (160 rpm). The transesterification capability of the crude extracellular lipase from LPB1 was assessed using thin layer chromatography by using hexane/diethyl ether/acetic acid as solvent system in the ratio of 90:10:1 (v/v/v). The free extracellular lipase exhibited the formation of methyl esters with the vegetable oils tested such as karanja (Rf 0.59), neem (Rf 0.59), castor (Rf 0.6) and olive oil (Rf 0.62). Both the soluble and immobilized lipase of this isolate demonstrated the methanolysis of non edible oil of Karanja (Pongamia pinnata) within 1-3 h.
PDF Abstract XML References Citation
Received: April 09, 2010;
Accepted: October 13, 2010;
Published: November 04, 2010
How to cite this article
P. Gupta, L.S.B. Upadhyay and R. Shrivastava, 2011. Lipase Catalyzed-transesterification of Vegetable Oils by Lipolytic Bacteria. Research Journal of Microbiology, 6: 281-288.
URL: https://scialert.net/abstract/?doi=jm.2011.281.288
URL: https://scialert.net/abstract/?doi=jm.2011.281.288
INTRODUCTION
Lipases (Triacylglycerol acylhydrolases EC 3.1.1.3) are a class of hydrolases which catalyze the hydrolysis of triglycerides to glycerol and free fatty acids on an oil-water interface (Kamini et al., 2000). In addition they are carboxylesterases that catalyze the hydrolysis and transesterification of esters. The syntheses of esters can exhibit enantioselective properties (Varma et al., 2007; Bezbradica et al., 2007; Abbas et al., 2002). Apart from its prevalent use in biosurfactants, aroma and flavor compounds, lubricant and in polyesters, it has also find use in biodiesel preparation (Edmundo et al., 1998; Athawale et al., 2003; Hills, 2003; Kumar and Gross, 2000; Jaeger and Eggert, 2002). Lipase has been found in many species like Gryllus campestris (Ozlem et al., 2007) and Cocos nucifera Linn. (Ejedegba et al., 2007). Lipolytic bacteria can be isolated from soil, raw milk and waste water (Mohan et al., 2008; Abdou et al., 2003; Bhumibhamon et al., 2002). Several bacterial species producing extracellular lipases have been reported (Bora and Kalita, 2009; Amoozegar et al., 2008; Zeng et al., 2004). Effective transesterification reactions using s everal lipases from P. aeruginosa MTCC 5113, E. aerogenes, P. fluroscens MTCC 103, have been also reported by several researchers (Singh et al., 2008; Kumari et al., 2009; Devanesan et al., 2007).
Transesterification of vegetable oils received considerable attention in past few years, which produces fatty acid and alkyl esters that are valuable intermediates in oleo chemistry and ethyl esters which are excellent substitute for diesel fuels (Freedman et al., 1986; Schuchardt et al., 1998). Enzymes perform very specific transesterification reaction (bio-transformations) (Stamatis et al., 2000); have made them increasingly popular in industries where less specific chemical processes produce unwanted by-products. The disadvantage associated with enzymatic transesterification is cost of enzyme preparation. Immobilization generally increases the reusability of the enzyme (Roy et al., 2003) and hence lowers the cost as well as helps the biocatalyst to become efficient in nonaquous media (Shah et al., 2007).
Furthermore, more than hundred types of tropical plants that produce oil-bearing materials have been identified. Among them are rubber seed oil, fenugreek oil, Neem oil, karanja oil, castor oil, olive oil and jatropha oil. They are good sources of oils with special fatty acid compositions and therefore have to be exploited by much research as excellent substrate for transesterification reaction (Shah et al., 2004; Kumari et al., 2009).
The present study reports the isolation of lipase producing bacteria from soil and to assess both the free and immobilized lipase preparations for transesterificaion of vegetable oils in a solvent-free system to produce methyl esters.
MATERIALS AND METHODS
Sample collection: Soil samples were aseptically collected from Raipur city with the help of soil auger in sterile sealed plastic bags for isolation of lipase producing bacteria under laboratory conditions in the year 2009. The soil samples from oil spilled areas included Kitchen garden waste, dairy farm compost (Sanchi dairy industry), compost yard (Kedia distillery limited) and Jatropha oil mill waste.
Isolation of lipolytic bacteria: The lipolytic bacteria were isolated from collected soil samples by serial dilution method. For this 1.0 g of soil sample was dissolved in 100 mL of sterile distilled water. This was serially diluted (10-1 to 10-5) and diluted samples were plated on solid agar medium containing olive oil in enrichment media (0.5% w/v (NH4)2 SO4, 0.05% w/v K2HPO4, 0.03% w/v MgSO4.7H2O, 2% v/v olive oil and 1.5% w/v agar, pH 7.0). Colony capable for utilizing olive oil as sole source of carbon was isolated and individually streaked on modified lipase assay media containing 1.5% w/v peptone, 0.5% w/v NaCl, 0.1% w/v CaCl2, 1% v/v Tween 80, 1.5% w/v agar at pH 7.2 as described by Shukla and Gupta (2007). The plates were incubated at 30°C for 72 h. The formation of white precipitate of calcium monostearate around the colony was considered as positive colony for extracellular lipase secretion and used for further study. Gram staining was carried out as given by Brucker (1986). The stock cultures were maintained routinely on nutrient agar media.
Lipase production: The pure culture of lipolytic bacteria was maintained on nutrient agar medium (0.5% w/v peptone, 5% w/v beef extract, 0.15% w/v yeast extract, 0.5% w/v NaCl and 1.5% w/v agar pH 7.5). The extra cellular lipase production was carried out in a medium composed of 3% w/v yeast extract, 1% w/v KH2PO4, 0.1% w/v MgSO4.7H2O, 0.5% w/v maltose and 0.2% v/v olive oil at pH 7.2. Medium was sterilized and inoculated with 1% seed inoculum prepared in nutrient broth followed by incubation at 37°C for 48 h at 160 rpm in a shaker incubator. By using different substrate sources such as olive oil, olive oil and lecithin (emulsifier), tween 80 and castor oil, their effect on lipase production by selected bacterial isolate was assessed at pH of 7.2. The cells were then centrifuged at 10,000 rpm for 15 min and the supernatant was used directly as crude preparation of lipase for further studies.
Lipase assay: The lipase activity in the culture supernatant was determined by titrimetric method (Sadasivam and Manickam, 1991). Lipase activity (E.C. 3.1.1.3) was measured by titration the fatty acid released with 0.1 M NaOH using 0.1% alcoholic phenolphthalein as indicator.
One unit of lipase activity was defined as the amount of enzyme releasing one mole of free fatty acid in 1 min under standard assay condition.
Immobilization of extracellular lipase by entrapment: The sodium alginate entrapment of crude lipase was carried out according to the standard method (Bhushan et al., 2008). Alginate with a concentration range of 4-10 % and LB1 lipase preparation were mixed together and stirred for 15 min at 4°C to obtain a homogenous mixture. Alginate-lipase homogenous mixture was extruded drop by drop into a cold CaCl2 solution (0.4 M) through pipette. The beads were stirred in CaCl2 solution for 45 min and then thoroughly washed with buffer (Tris acetate 0.107 M, pH 7.2). Alginate immobilized lipase beads were stored at 4°C in Tris acetate buffer till further use.
Set up for transesterification reaction: Transesterification reaction was carried out for different oil samples such as Olive, Neem, Karanja and Castor oil separately with a short chain alcohol, i.e., methanol. Oil and methanol were taken in the ratio of 1:4 (mol/mol) in a screw capped vial. To this oil: methanol mixture 2 mL of crude lipase was added and incubated at 40°C with constant stirring at 200 rpm for 3 h in a shaker incubator. Process of methanolysis was performed with both free and immobilized lipase preparation.
Analysis of esters: The formation of methyl esters of vegetable oil in the reaction mixture was analyzed by thin-layer chromatography on silica gel H (E. Merck, Mumbai, India) plates. The coated silica gel plates were spotted with transesterified oil samples and bio-diesel. The chromatogram was developed in chromatographic chamber using hexane/diethyl ether/acetic acid as solvent system in the ratio of 90:10:1 (v/v/v). The spots were detected in the iodine chamber and Rf values were calculated and compared with authentic standards (Gordon et al., 1994).
RESULT AND DISCUSSION
Isolation and screening of lipolytic bacteria: The lipase producing bacteria were isolated from different soil samples. Enrichment culture technique and lipase assay media enable the isolation of extracellular lipase producing bacteria. In total, 15 isolates were isolated by enrichment culture technique from the soil samples and among them; three isolates (LPB1, LPB4 and LPB7) showed measurable lipolytic activity as shown in Fig. 1a-d. The precipitation zone for strain LPB1 (Fig. 1) was found to be appreciable among the three isolates and hence was further explored for lipase production and transesterification process. The LPB1 was identified as gram negative and rod shaped on gram staining.
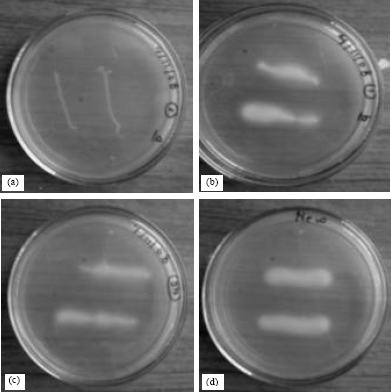 | |
| Fig. 1: | Photo images of zone of precipitation generated by isolates of lipase producing bacteria on Tween 80 agar medium. (a) LPB12, negative isolate from compost yard the positive isolates (b) LPB7, (c) LPB4 and (d) LPB1 from kitchen garden soil, Jatropha oil mill waste and dairy farm compost, respectively |
Lipase production: The lipase production efficiency of lipolytic bacteria was assayed in presence of different substrates i.e., Olive oil, Olive oil and Lecithin (emulsifier), Castor oil and Tween 80. The assay was also performed at various pH ranges i.e., from 5.6 to 8.0. Optimization of extracellular lipase production was carried out in bacterial (Babu and Rao, 2007) and fungal (Iftikhar and Hussain, 2002) lipase. It was observed that strain LPB1 produces lipase at all pH range tested. With an increase in the pH from 5.6 to 7.2 the lipase production was increased in isolate LPB1 from 0.006 to 0.0077 (μg/mL/min). Beyond pH 7.2 a fall in lipase production was observed (Fig. 2). Out of the six different values of pH of production medium the pH optima for maximum production of lipase in strain LPB1 was found to be 7.2 (0.0099 μg/mL/min). The production of lipase decreased in slightly acidic (pH 5.6) or alkaline (pH 8.0) whereas, at pH 7.2 the highest production was reported. The production of extracellular lipase can be induced by using the lipidic substrate (Akhtar et al., 1980). Hence, the lipase production was found measurable for all tested lipidic substrate (Fig. 3). Olive oil was found to be the best substrate for lipase production among the substrates tested. The lipase activity was found comparable in presence of Olive oil and Tween 80 (0.0070 μg/mL/min and 0.0077 μg/mL/min, respectively). An increase in lipase activity was observed from 0.007 to 0.009 (μg/mL/min) in presence of lecithin as emulsifier in production media containing Olive oil as substrate at pH 7.2. The incorporation of surface active substances can increase the availability of lipidic substrate by emulsifying the lipid and could further increase the extracellular lipase activity (Naka et al., 1986). The value reported by us are almost similar to that obtained for lipase production from Bacillus sp. (Mohan et al., 2008; Achamma et al., 2003). Achamma et al. (2003) has also reported maximum production of lipase when olive oil has been used as substrate.
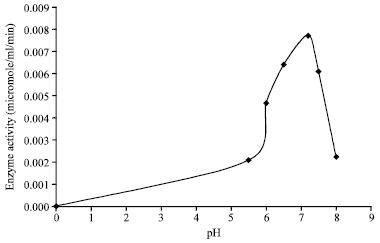 | |
| Fig. 2: | Effect of pH on lipase production by isolate LPB1 in production media. The result is a mean of four observations |
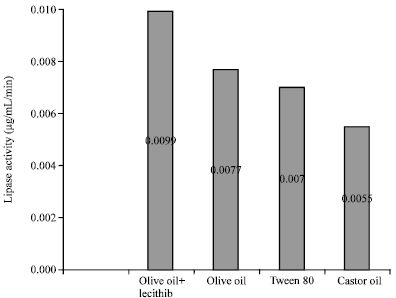 | |
| Fig. 3: | Effect of various substrates on lipase production by isolate LPB1 at pH 7.2. The result is a mean of four repeats |
Similar result indicating olive oil to be the best substrate of lipase production in various strains of fungi has been reported by Annibale et al. (2006).
Transesterification: Formation of methyl ester of various oil samples (Karanja, Castor, Neem and Olive) has been monitored by thin layer chromatography on silica gel plate employing hexane/diethyl ether/acetic acid as solvent system. The spots were detected in the iodine chamber and identified by comparison with the Rf values of authentic standards form literature for methyl oleate (Gordon et al., 1994). Figure 4a shows the methyl ester spots obtain and developed by TLC of oil samples transesterified by lipase obtained from LPB1. Transesterification of different oil samples such as karanja (Rf 0.59), neem (Rf 0.59), castor (Rf 0.6) and olive oil (Rf 0.62), respectively could be confirmed since the Rf values of samples calculated is comparable with the Rf value of standard methyl oleate ester (Rf 0.63).
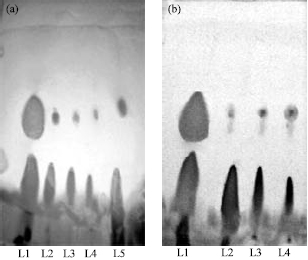 | |
| Fig. 4: | (a) Standard methyl oleate and methyl esters of oil samples tested when observed from left to right in the silica gel TLC plates after transesterification reaction (Solvent system: hexane/diethyl ether/acetic acid, 90:10:1 (v/v/v)). TLC of methanolyzed samples using crude LPB1 lipase. Lanes1, Standard methyl esters (Rf 0.63), lanes 2-5 reaction mixture of Karanja oil (Rf 0.59), Neem oil (Rf 0.59), Castor Oil (Rf 0.6), Olive oil (0.62) (Left to right), (b) TLC analysis of reaction mixture after methanolysis of karanja oil using immobilized LPB1 lipase. Lanes1, Standard methyl esters (Rf 0.63), Lanes 2-4 reaction mixture of Karanja oil after 1-3 hr (Rf 0.6, 0.6, 0.61) |
Hence we report the methonlysis of oil samples catalyzed by LPB1 crude lipase isolated from dairy compost (Sanchi, Raipur).
Karanja oil has been chosen for the time dependent tranesterification studies since it is cheap, non edible and present abundantly in India. Hence, it can serve as a good source of raw material for bio fuel generation. Since, immobilization impart stability to enzyme system and also increases the shelf life and reusability of enzyme. Hence an attempt has been made to check out the efficiency of LPB1 lipase in immobilized condition for transesterification reaction. Figure 4b shows the transesterification of Karanja oil at various interval of time (1-3 h) by alginate immobilized LPB1 crude lipase. Transesterification started within first hours of incubation of Karanja oil with immobilized lipase. Transesterification has been worked out in aqueous and non aqueous system using lipase for bio-diesel production by many workers (Singh et al., 2008). In order to utilize lipases in non aqueous environment the enzyme has frequently been immobilized on various support matrix (Jagannathan, et al., 2008; Shieh et al., 2003; Hus et al., 2002). Alginate has been used as immobilization matrix for lipase immobilization by many workers (Bhushan et al., 2008; Devanesan et al., 2007).
From the present study it has been demonstrated that the LPB1 strain shows the efficiency of methanolysis of non edible oil from Karanja. The crude lipase of LPB1 both in soluble form as well as alginate immobilized form exhibited the transesterification of this oil within 3 h of incubation of reaction mixture. In comparison with the highly purified commercial lipase, the inexpensive crude lipase preparation obtained from LPB1 had a distinct rate of tranesterification and at the same time is comparatively cheaper. However,,the further research on various parameters affecting the methyl ester yields namely temperature, pH, reaction time and number of bead and molecular ratio of oil to methanol is to be studied.
REFERENCES
- Abbas, H., A. Hiol, V. Deyris and L. Comeau, 2002. Isolation and characterization of an extracellular lipase from Mucor sp. strain isolated from palm fruit. Enzyme Microbial Technol., 31: 968-975.
CrossRef - Akhtar, M.W., A.Q. Miraz and M.D.I. Chughtai, 1980. Lipase induction in Mucor hiemalis. Applied Environ. Microbiol., 18: 257-263.
Direct Link - Amoozegar, M.A., E. Salehghamari, K. Khajeh, M. Kabiri and S. Naddaf, 2008. Production of extracellular thermohalophilic lipase from a moderately halophilic bacterium, Salinivibrio sp. Strain SA-2. J. Basic Microbiol., 48: 160-167.
PubMed - Kumari, A., P. Mahapatra, V.K. Garlapati and R. Banerjee, 2009. Enzymatic transesterification of Jatropha oil. Biotechnol. Biofuels, 2: 1-7.
CrossRefPubMedDirect Link - Athawale, V., N. Manjrekar and M. Athawale, 2003. Effect of reaction parameter on synthesis of citronellyl methacrylate by lipase-catalyzed transesterification. Biotechnol. Prog., 19: 298-302.
PubMed - Bezbradica, D., D. Mijin, S. Siler-Marinkovic and Z. Knezevic, 2007. The effect of substrate polarity on the lipase-catalyzed synthesis of aroma esters in solvent-free systems. J. Mol. Catalysis B. Enzyme, 45: 97-101.
CrossRef - Bhumibhamon, O., K. Achara and F. Suptawee, 2002. Biotreatment of high fat and oil wastewater by lipase producing microorganisms. Kasetsart J. Nat. Sci., 36: 261-267.
Direct Link - Bhushan, I., R. Parshad, G.N. Qazi and V.K. Gupta, 2008. Immobilization of lipase by entrapment in Ca-alginate beads. J. Bioactive Compatible Polym., 23: 552-562.
CrossRefDirect Link - Brucker, M.C., 1986. Gram staining a useful laboratory technique. J. Nurse-Midwifery, 31: 156-158.
PubMed - Devanesan, N.G., T. Viruthagiri and N. Sugumar, 2007. Transesterification of jatropha oil using immobilized Pseudomonas fluorescens. Afr. J. Biotechnol., 6: 2497-2501.
Direct Link - Edmundo, C., D. Valerie, C. Didier and M. Alain, 1998. Efficient lipase-catalyzed production of tailor-made emulsifier using solvent engineering coupled to extractive processing. J. Am. Chem. Soc., 75: 309-313.
CrossRef - Ejedegba, B.O., E.C. Onyeneke and P.O. Oviasogie, 2007. Characteristics of lipase isolated from coconut (Cocos nucifera Linn) seed under different nutrient treatments. Afr. J. Biotechnol., 6: 723-727.
Direct Link - Gordon, J.A., S.K. Heller, T.L. Kaduce and A.A. Spector, 1994. Formation and release of a peroxisome-dependent arachidonic acid metabolite by human skin fibroblasts. J. Biol. Chem., 269: 4103-4109.
PubMed - Hills, G., 2003. Industrial use of lipases to produce fatty acid esters. Eur. J. Lipid Sci. Technol., 105: 601-607.
Direct Link - Iftikhar, T. and A. Hussain, 2002. Effect of nutrients on extracellular lipase production by mutant strain of Rhizopus oligosporus Tuv-31. Biotechnology, 1: 15-20.
Direct Link - Jaeger, K.E. and T. Eggert, 2002. Lipases for biotechnology. Curr. Opin. Biotechnol., 13: 390-397.
CrossRefPubMedDirect Link - Kumar, A. and R.A. Gross, 2000. Candida antarctica lipase B catalyzed transesterification: New synthetic routes to copolymers. J. Am. Chem. Soc., 122: 11767-11770.
CrossRef - Roy, I., A. Gupta, S.K. Khare, V.S. Bisaria and M.N. Gupta, 2003. Immobilization of xylan-degrading enzymes from Melanocarpus albomyces IIS 68 on the smart polymer Eudragit L-100. Applied Microbiol. Biotechnol., 61: 309-313.
CrossRef - Schuchardt, U., R. Sercheli and R.M. Vargas, 1998. Transesterfication of vegetable oils: A review. J. Braz. Chem. Soc., 9: 199-210.
CrossRef - Mohan, T.S., A. Palavesam and G. Immanvel, 2008. Isolation and characterization of lipase-producing Bacillus strains from oil mill waste. Afr. J. Biotechnol., 7: 2728-2735.
Direct Link - Shah, S., S. Sharma and M.N. Gupta, 2004. Biodiesel preparation by lipase catalyzed transesterification of jatropha oil. Energy Fuels, 18: 154-159.
CrossRef - Shah, S., K. Solanki and M.N. Gupta, 2007. Enhancement of lipase activity in nonaqueous media upon immobilization on multiwalled carbon nanotubes. Chem. Central J., 1: 30-30.
Direct Link - Stamatis, H., E.C. Voutsas, C. Delimitsou, F.N. Kolisis and D. Tassios, 2000. Enzymatic production of alkyl esters through lipase cataysed transesterification reaction in organic solvents: Solvent effects and prediction capabilities of equilibrium conversions. Biocatalysis Biotransformations, 18: 259-269.
Direct Link - Singh, M., S. Singh, R.S. Singh, Y. Chisti and U.C. Banerjee, 2008. Transesterification of primary and secondary alcohols using Pseudomonas aeruginosa lipase. Bioresour. Technol., 99: 2116-2120.
CrossRef - Varma, R., S.M. Kasture, B.G. Gaikwad, S. Nene and U.R. Kalkote, 2007. Lipase catalysed ennantioselective amidation of α phenyl-ethylamine. Asian J. Biochem., 2: 279-283.
Direct Link - Zeng, Z., X. Xiao, P. Wang and F. Wang, 2004. Screening and characterization of psychrotrophic, lipolytic bacteria from deep-sea sediments. J. Microbiol. Biotechnol., 14: 952-958.
Direct Link - Bora, L. and M. Kalita, 2009. Production of extracellular lipase from Bacillus sp. LBN4 by solid state fermentation. Internet J. Bioeng., Vol. 3, No. 2.
Direct Link - Abdou, A.M., 2003. Purification and partial characterization of psychrotrophic Serratia marcescens lipase. J. Dairy Sci., 86: 127-132.
CrossRefDirect Link - Shukla, P. and K. Gupta, 2007. Ecological screening for lipolytic molds and process optimization for lipase production from Rhizopus Oryzae KG-5. J. Applied Sci. Environ. Sanitation, 2: 35-42.
Direct Link - Shieh, C.J., H.F. Liao and C.C. Lee, 2003. Optimization of lipase-catalyzed biodiesel by response surface methodology. Bioresour. Technol., 88: 103-106.
CrossRefDirect Link - D'Annibale, A., G.G. Sermanni, F. Federici and M. Petruccioli, 2006. Olive-mill wastewaters: A promising substrate for microbial lipase production. Bioresour. Technol., 97: 1828-1833.
CrossRef
Balaji M. Panchal Reply
Very good research paper in 2011 year