
Gang Xie
Department of Chemistry and Biochemistry,
South Dakota State University, Brookings, SD 57007, USA
Thomas P. West
Department of Biology and Microbiology,
South Dakota State University, Brookings, SD 57007, USA
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 9 | Page No.: 678-683







ABSTRACT
Seven strains of the fungus Aspergillus niger were screened for their ability to produce citric acid on the ethanol dry milling coproduct thin stillage. Citric acid and biomass production by the fungal strains grown on the thin stillage were analyzed using an enzyme assay and a gravimetric method, respectively. Citric acid production by A. niger ATCC 9029, ATCC 9142, ATCC 10577, ATCC 12846, ATCC 26550 and ATCC 201122 was similar after 144 h of growth on thin stillage. These strains also exhibited high citric acid specific productivities and yields. Only the citric acid production, specific productivity and yield by A. niger ATCC 11414 were lower after growth on thin stillage. Biomass production by ATCC 9029 on thin stillage was the highest of the strains studied. For all strains, at least 88% of the reducing sugar concentration in the thin stillage was consumed after 144 h.
PDF Abstract XML References
How to cite this article
Gang Xie and Thomas P. West, 2007. Citric Acid Production by Aspergillus niger on the Ethanol Dry Milling Coproduct Thin Stillage. Research Journal of Microbiology, 2: 678-683.
URL: https://scialert.net/abstract/?doi=jm.2007.678.683
URL: https://scialert.net/abstract/?doi=jm.2007.678.683
INTRODUCTION
Citric acid is a commercially important specialty chemical that has a number of industrial applications for use in beverages, foods and pharmaceuticals (Tran et al., 1998). Some applications of citric acid include utilization as a preservative, flavor enhancer, a chelating agent, pH regulator, antioxidant and stabilizer. Global production of citric acid is estimated to be over 1.4 million tons (Soccol et al., 2006). Strains of the fungus Aspergillus niger have been shown to excrete citric acid (Legisa and Mattey, 2007) and also have been tested for their ability to produce citric acid from brewery wastes (Hang et al., 1975, 1977; Roukas and Kotzekidou, 1986). Other substrates that have been utilized for citric acid fermentation by A. niger were beet molasses, carobod syrup or corn starch (Clement, 1952; Macris, 1975; Nguyen et al., 1992). It was of interest to learn whether the coproduct thin stillage resulting from the ethanol dry milling process could be utilized as a substrate for citric acid production by A. niger strains. The major coproducts produced during ethanol production from corn using dry milling are corn distillers grains and thin stillage. Previous studies have shown that citric acid can be produced by A. niger strains grown on corn distillers grains (Xie and West, 2006a-c). The thin stillage is recovered from the whole stillage after centrifugation. The thin stillage is mixed with the wet corn distillers grains and dried to produce corn distillers grains with solubles. About 18 pounds of 90% corn distillers grains with solubles are produced from each bushel of corn processed at dry milling ethanol plants. Currently, corn distillers grains with solubles is used as a protein supplement in animal feeds (Ham et al., 1994). With more than a million tons of grains being produced from the ethanol dry milling process per year and the wet distillers grains also serving as an animal feed, the low value thin stillage could be more fully utilized to produce a specialty chemical such as citric acid. In this study, seven strains of A. niger were screened for their ability to produce citric acid from thin stillage by surface fermentation.
MATERIALS AND METHODS
Strains and Growth Conditions
Seven citric acid-producing strains, namely Aspergillus niger ATCC 9029 (Somkuti and Bencivengo, 1981), ATCC 9142 (Doelger and Prescott, 1934), ATCC 10577 (Clement, 1952), ATCC 11414 (Perlman et al., 1946), ATCC 12846 (Moyer, 1953), ATCC 26550 (Martin and Waters, 1952) and ATCC 201122 (Gradisnik-Grapulin and Legisa, 1996) were used in this study. The seven strains were tested for their ability to produce citric acid from the thin stillage using surface fermentation. The experiments were conducted (Brookings, SD, USA) in August 2005 and the source of the thin stillage was Dakota Ethanol LLC (Wentworth, SD, USA). The thin stillage was prepared as a substrate by initially adjusting its pH to 6.0 and filtering it through a Whatman No. 1 filter to remove insoluble material. The clarified stillage was sterilized at 117.2 kPa of pressure for 20 min at 121°C. A volume of the processed thin stillage was added to sterile 250 mL Erlenmeyer flasks. To prepare the inoculum, a loopful of fungal mycelium of the A. niger strain was inoculated into potato dextrose broth (20 mL) and the culture was grown for 72 h at 25°C. A 5% inoculum (2x103 conidia mL-1) of each strain was added to each flask to initiate the surface fermentation of the thin stillage. The cultures were placed in a rotary shaker and shaken at 200 rpm at 25°C for a period of 144 h.
Processing of Grains
Following the incubation period of 144 h, the cultures were processed to allow the assaying of citric acid and biomass levels. Each culture was filtered through a Whatman No. 1 filter. The fungal biomass in each culture was washed with sterile water (10 mL) and also filtered through a Whatman No. 1 filter. The fungal biomass was removed and utilized during the subsequent biomass determination. To precipitate any protein present in each culture filtrate, ice cold 0.5 M HClO4 (0.5 mL) was added and the filtrate was stirred. After any protein precipitate present was removed by filtration, the filtrate was subsequently neutralized. The volume of each culture filtrate was recorded.
Citric Acid Assay
The citric acid content of the neutralized filtrate was assayed spectrophotometrically using a coupled enzyme assay (Moellering and Gruber, 1966; Henniger and Mascaro, 1985). The composition of the modified assay mix (1 mL) was 0.1 M glycylglycine buffer pH 7.8, 0.2 mM NADH, 0.6 mM ZnCl2, 5 units citrate lyase, 6 units malate dehydrogenase, 3 units lactate dehydrogenase and sample. Standard concentrations of citric acid were also assayed. The reaction was followed at 340 nm by monitoring the decrease in absorbance that is proportional to the concentration of citric acid present in the sample. Citric acid levels are expressed as g citric acid L-1 thin stillage. All values represent the mean of three separate determinations involving three independent cultures. The Student’s t-test was utilized during statistical analysis.
Biomass and Reducing Sugar Determinations
To determine biomass production, wet fungal biomass collected after 144 h of growth was placed in a preweighed beaker and dried at 105°C to constant weight. The beaker containing the dry fungal biomass was reweighed to derive the weights for each culture. The weight of the inoculum added to each culture was determined by collection on preweighed filters, drying to constant weight at 105°C and subtracting these weights from the biomass levels determined after 144 h. Reducing sugar levels were determined using a previously described assay where glucose served as the standard (Dygert et al., 1965). All values represent the mean of three independent determinations involving three separate cultures. The Student’s t-test was used during statistical analysis.
RESULTS AND DISCUSSION
It has previously been shown that strains of Aspergillus niger excrete citric acid after growth on corn substrates such as ground corn and corn starch (Moyer, 1953; Nguyen et al., 1992). It has also been shown that citric acid can be produced by Aspergillus niger strains grown on brewery wastes (Hang et al., 1975, 1977; Roukas and Kotzekidou, 1986). In this study, it was shown that seven citric acid-producing strains of A. niger were able to utilize stillage for citric acid production after 144 h at 25°C (Table 1). Six of the seven strains, namely ATCC 9029, ATCC 9142, ATCC 10577, ATCC 12846, ATCC 26550 and ATCC 201122, were capable of producing similar citric acid levels after 144 h of growth on thin stillage (Table 1). No statistically significant difference in the citric acid levels produced by these six strains was observed. There was a statistically significant (p<0.01) difference in the levels of citric acid produced by these strains compared to ATCC 11414. ATCC 11414 produced the lowest level of citric acid following its growth on thin stillage for 144 h (Table 1). With respect to biomass production, ATCC 9029 produced the highest biomass level on the thin stillage after 144 h of growth (Table 1). The difference in biomass production by this strain was statistically significant compared to biomass production by ATCC 9142 (p<0.05), ATCC 10577 (p<0.05), ATCC 11414 (p<0.05), ATCC 12846 (p<0.01), ATCC 26550 (p<0.01) and ATCC 201122 (p<0.01). The specific productivities of ATCC 9029, ATCC 9142, ATCC 10577, ATCC 12846, ATCC 26550 and ATCC 201122 were comparable with the lowest specific productivity being noted for ATCC 11414 (Table 2). A statistically significant difference between their specific productivities was not observed. The citric acid yields produced by ATCC 9142, ATCC 10577, ATCC 11414, ATCC 12846, ATCC 26550 and ATCC 201122 were similar (Table 2). The citric acid yield produced by ATCC 11414 was found to be statistically lower (p<0.01) than the other strains studied (Table 2). Relative to reducing sugar consumption by the strains, it can be noted that a high reducing sugar concentration is utilized by all seven strains (Table 3). This is clearly evident because nearly 90% of the available reducing sugar concentration of the thin stillage is consumed by all the strains tested (Table 3).
| Table 1: | Citric acid and biomass production by Aspergillus niger strains grown on thin stillage as a substrate for 144 h at 25°C |
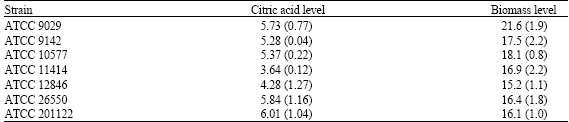 | |
| The citric acid levels are expressed as g citric acid L-1 thin stillage while the biomass levels are given as g cell weight L-1 thin stillage. All values represent the mean of three separate trials (standard deviation) | |
| Table 2: | Citric acid specific productivity and yield by Aspergillus niger strains grown on thin stillage as a substrate |
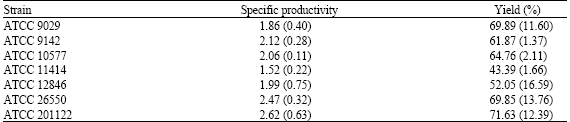 | |
| Specific productivity is expressed as g citric acid kg-1 biomass h-1 while % yield is given as g citric acid g-1 reducing sugar. All values represent the mean of three separate trials (standard deviation) | |
| Table 3: | Reducing sugars consumed by Aspergillus niger strains following growth on thin stillage as a substrate for 144 h at 25°C |
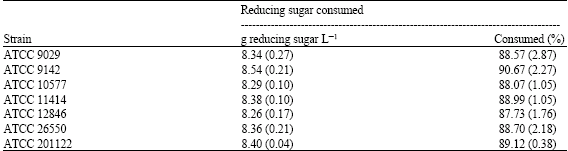 | |
| The results indicate the mean of three separate trials (standard deviation) | |
Although limited in number, prior studies have examined citric acid production by A. niger using surface fermentation of brewery wastes (Hang et al., 1975, 1977; Roukas and Kotzekidou, 1986). Using lager tank sediment as a substrate, A. niger ATCC 10577 produced about 7-8 g L-1 citric acid on spent grain liquor after 192 h of growth at 30°C (Roukas and Kotzekidou, 1986). An initial study found that Aspergillus foetidus ATCC 10254 produced 8 g L-1 citric acid and 12 g L-1 dry mycelial weight of growth on spent grain liquor after 96 h at 30°C (Hang et al., 1975). A more comprehensive study showed that A. foetidus ATCC 10254 produced an average of 7.6 g L-1 citric acid with an average citric acid yield of 49.9% when it was grown on spent grain liquor for 96 h at 30°C although some brewery waste samples only supported 3.5 g L-1 citric acid (Hang et al., 1977). The reducing sugar concentration of the spent grain liquor consumed by the fungus averaged about 86% (Hang et al., 1977). The mycelial dry weight ranged from 6-11 g L-1 after growth for 96 h at 30°C (Hang et al., 1977). Brewery wastes were reported to be excellent substrates for biomass production by yeast and fungal strains (Shannon and Stevenson, 1975). Similar to this investigation, some of the same strains used in this work were utilized in the previous studies and all the previous reports employed surface fermentation (Hang et al., 1975, 1977; Roukas and Kotzekidou, 1986). This study was also similar to prior reports in that the fermentation wastes used contained comparable initial reducing sugar concentrations (Hang et al., 1975, 1977). There was only one study that differed from this study in that the fermentation waste contained a higher initial level of reducing sugars (Roukas and Kotzekidou, 1986). A comparison of the findings of this investigation with the earlier studies indicated that similar levels of citric acid could be produced by the majority of the A. niger strains tested on the fermentation wastes (Hang et al., 1975, 1977). Biomass production by ATCC 9142 and ATCC 10577 was higher after growth on thin stillage compared to their biomass production on other fermentation wastes (Hang et al., 1975, 1977; Roukas and Kotzekidou, 1986) which is likely due to the higher protein content in the thin stillage supporting better fungal growth. Further, the citric acid yield observed for the majority of strains used in this study was higher than 60% and this compared well with the data from prior investigations (Hang et al., 1975, 1977). It is clear that thin stillage could be used by all of the A. niger strains screened as a substrate for the production of citric acid and supported high levels of biomass production. The percentage of reducing sugar consumed by the A. niger strains tested in this study was nearly equivalent to the percentage observed when A. foetidus was grown on spent grain liquor (Hang et al., 1977).
CONCLUSIONS
Citric acid was produced by all the A. niger strains grown on thin stillage at comparable levels for six of the seven strains tested. Moreover, the citric acid specific productivities and yields were highest for these six strains. Biomass production on thin stillage was highest for A. niger ATCC 9029 but the thin stillage supported excellent growth by all the strains tested. Compared to other brewery wastes, thin stillage appeared to be as effective a substrate for fungal citric acid production.
ACKNOWLEDGMENTS
Published as paper 3526, Journal Series, South Dakota Agricultural Experiment Station. This research was supported by the South Dakota AES and South Dakota Corn Utilization Council. This manuscript reports results of research only and the mention of brand or firm names does not constitute an endorsement by the South Dakota AES over those mentioned but of a similar nature.
REFERENCES
- Doelger, W.P. and S.C. Prescott, 1934. Citric acid fermentation. Ind. Eng. Chem., 26: 1142-1149.
CrossRefDirect Link - Dygert, S., L.H. Li, D. Florida and J.A.Thoma, 1965. Determination of reducing sugar with increased precision. Anal. Biochem., 13: 367-374.
Direct Link - Gradisnik-Grapulin, M. and M. Legisa, 1996. Comparison of specific metabolic characteristics playing a role in citric acid excretion between some strains of the genus Aspergillus. J. Biotechnol., 45: 265-270.
CrossRef - Ham, G.A., R.A. Stock, T.J. Klopfenstein, E.M. Larson, D.H. Shain and R.P. Huffman, 1994. Wet distillers byproducts compared with dried distillers grains with solubles as a source of protein and energy for ruminants. J. Anim. Sci., 72: 3246-3257.
Direct Link - Hang, Y.D., D.F. Splittstoesser and E.E. Woodams, 1975. Utilization of brewery spent grain liquor by Aspergillus niger. Applied Microbiol., 30: 879-880.
Direct Link - Hang, Y.D., D.F. Splittstoesser, E.E. Woodams and R.M. Sherman, 1977. Citric acid fermentation of brewery waste. J. Food Sci., 42: 383-384.
CrossRef - Henniger, G. and L. Mascaro, Jr., 1985. Enzymatic-ultraviolet determination of L-citric acid in wine collaborative study. J. Assoc. Off. Anal. Chem., 68: 1024-1027.
Direct Link - Legisa, M. and M. Mattey, 2007. Changes in primary metabolism leading to citric acid overflow in Aspergillus niger. Biotechnol. Lett., 29: 181-190.
CrossRefDirect Link - Moellering, H. and W. Gruber, 1966. Determination of citrate with citrate lyase. Anal. Biochem., 17: 369-376.
CrossRefDirect Link - Moyer, A.J., 1953. Effect of alcohols on the mycological production of citric acid in surface and submerged culture. II. Fermentation of crude carbohydrates. Applied Microbiol., 1: 7-13.
Direct Link - Nguyen, T.K., L. Martinkova, L. Seichert and F. Machek, 1992. Citric acid production by Aspergillus niger using media containing low concentrations of glucose or corn starch. Folia Microbiol., 37: 433-441.
Direct Link - Roukas, T. and P. Kotzekidou, 1986. Production of citric acid from brewery wastes by surface fermentation using Aspergillus niger. J. Food Sci., 51: 225-228.
CrossRefDirect Link - Soccol, C.R., L.P.S. Vandenberghe, C. Rodrigues and A. Pandey, 2006. New perspectives for citric acid production and application. Food Technol. Biotechnol., 44: 141-149.
Direct Link - Tran, C.T., L.I. Sly and D.A. Mitchell, 1998. Selection of a strain of Aspergillus for the production of citric acid from pineapple waste in solid-state fermentation. World J. Microbiol. Biotechnol., 14: 399-404.
CrossRefDirect Link - Xie, G. and T.P. West, 2006. Comparison of citric acid production by Aspergillus niger ATCC 9029 and ATCC 12846 on corn distillers` grains with solubles. Res. J. Microbiol., 1: 540-545.
CrossRefDirect Link - Xie, G. and T.P. West, 2006. Citric acid production by Aspergillus niger on wet corn distillers grains. Lett. Applied Microbiol., 43: 269-273.
CrossRefPubMedDirect Link - Xie, G. and T.P. West, 2006. Citric acid production by Aspergillus niger on corn distillers` grains with solubles. Res. J. Microbiol., 1: 228-233.
CrossRefDirect Link