
Saira Anwar
Department of Botany, University of the Punjab, Quaid-e-Azam Campus, Lahore-54590, Pakistan
Anjum Nasim Sabri
Department of Botany, University of the Punjab, Quaid-e-Azam Campus, Lahore-54590, Pakistan
Shahida Hasnain
Department of Botany, University of the Punjab, Quaid-e-Azam Campus, Lahore-54590, Pakistan
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 66-73







ABSTRACT
The present study deals with the effect of temperature (37 and 45°C) in combination with varying pH (6, 7, 8, 9) after 24 and 96 h of incubation on cell morphology, staining behavior and growth rate of PY79 (wild type) and sporulation defective mutant strains (5, 19, 96, 99) of bacillus. Strains were isolated from local polluted area around Lahore. All the strains were gram positive spore former aerobic rods. In majority of strains the optimum temperature was 37°C and pH 7. Frequencies of different cell types differ at different temperatures and at varying pH. Cell degeneration and lysis was more in sporulation defective strains (5, 19, 96, 99). Spores which were formed by the bacterial isolates 5, 19, 96 and 99 were different from wild type and were unable to germinate.
PDF Abstract XML References
How to cite this article
Saira Anwar, Anjum Nasim Sabri and Shahida Hasnain, 2007. Study of Varying Temperature and pH on Bacterial Cell Morphology of Wild Type (PY79) and Sporulation Defective Strains of Bacillus. Research Journal of Microbiology, 2: 66-73.
URL: https://scialert.net/abstract/?doi=jm.2007.66.73
URL: https://scialert.net/abstract/?doi=jm.2007.66.73
INTRODUCTION
Bacillus subtilis is most extensively studied non pathogenic, aerobic, gram positive saprophytic rods found in soil. It belongs to industrially important group of organisms. There are over 150 genes involved in sporulation and germination that have been mapped on B. subtilis map (Dean and Zeigler, 1994). Sadaie and Kada (1985) have described the relationship between the initiation of cell division and initiation of sporulation, temperature, pH, starvation and enzyme action are the environmental factor which effects the cell division in B. subtilis (Chan et al., 1998; Helmann et al., 2001; Sabri and Hasnain, 1994, a, b; Kuile, 1994; Schut et al., 1995; Periago, 2002; Sachaik et al., 2004). In B. subtilis the non permissive temperatures involved the mutations and defects in cell division (Sato et al., 1984; Sadaie et al., 1991; Yanoun et al., 1993; Butler et al., 1993). pH response is important for growth and survival of bacteria in environment (Errington et al., 2003). It not only affect the morphogenetic processes in bacteria but also regenerates the expression of several genes (Castanie-Cornet and Foster, 2001; Stanick et al., 2002; Tucker et al., 2003). A major reason which may be involved in spore dormancy is the pH in the spore core, which is rather low, 6.3 for B. cerues spores, 6.4 for B. megaterium and B. subtilis was found to be ~ pH (Setlow and Setlow, 1980; Setlow and Johnson, 2002). The objective of the present study was to isolate and characterized bacillus strains and to compare them with wild type PY79.
MATERIALS AND METHODS
The present study was conducted in the Microbiology and Molecular Genetics Research Laboratory, Department of Microbiology and Molecular Genetics, University of the Punjab, Quaid-e-Azam Campus, Lahore-54590, Pakistan, in the year 2005.
| Table 1: | Strains used in the study |
 | |
Bacterial Strains
Bacterial strains used in this study were PY79, 5, 19, 96, 99 (Table 1).
Temperature and pH Range of Bacteria
The most suitable temperature for the growth of bacteria is 37°C at neutral pH 7. Eight test tubes of each bacterial strain were labeled for each temperature (37 and 45°C), pH (6, 7, 8, 9) and bacterial strains were taken in which 5 mL of nutrient broth were poured and then autoclaved. After autoclaving the 10 μL inoculums was given in each test tube from 24 h old fresh culture. The initial bacterial density (fixed at 0.5 optical density at 600 nm) was same for each strain.
Effect of Temperature and pH on Cell Morphology
Bacterial strains were incubated for different times (24, 96) at low different temperatures (37 and 45°C) at various pHs (6, 7, 8, 9). After gram staining the cell shapes and staining behavior was observed.
RESULTS
Gram Staining
All the strains used (PY79, 5, 19, 96, 99) were gram positive spore former rods.
Effect of Temperature and pH on Bacterial Growth
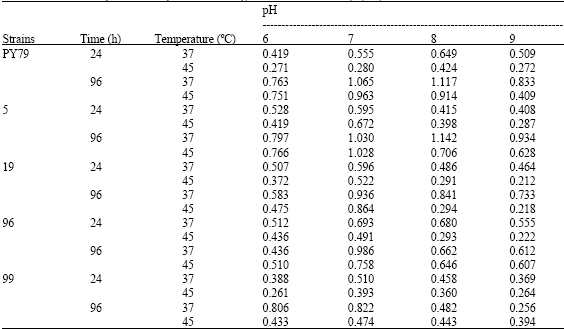
Bacterial isolates 19, 96, 99 preferred pH 7 (at 37 and 45°C) for maximal growth after 24 and 96 h. In strain PY79, after 24 and 96 h at both temperature (37 and 45°C) the optimum bacterial growth is recorded at pH 8.0 except after 96 h at 45°C the maximum growth is observed at pH 7 while in strain 5, after 24 and 96 h at different temperatures (37 and 45°C) at pH 7 there was maximum bacterial growth while after 96 h at temperature 37°C the bacterial growth is maximum at pH 8 (Table 2).
Effect of Temperature and pH on Cell Morphology
Strain-PY79
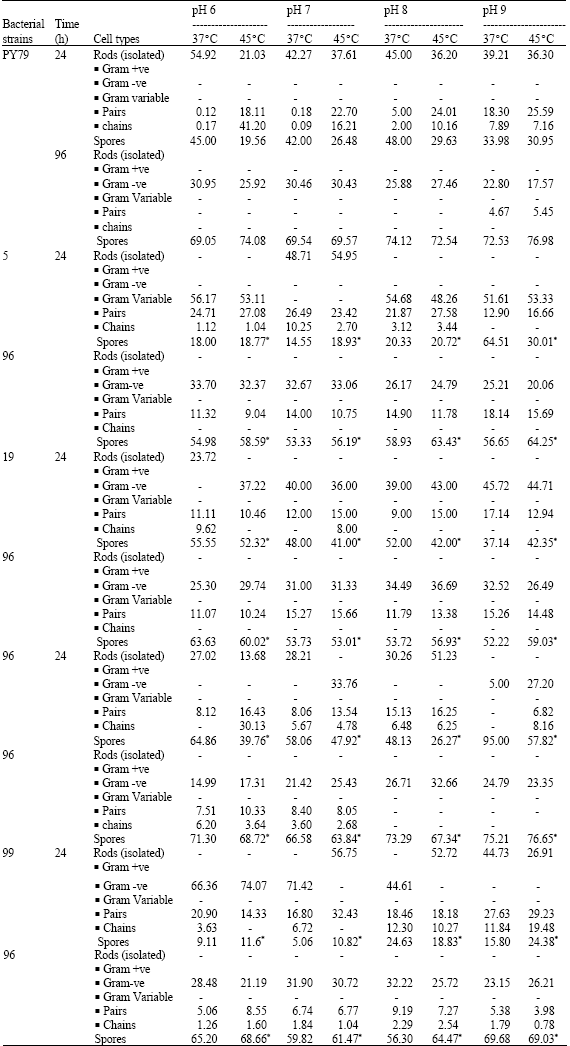
Alteration in the pH in combination of high temperature affected cell morphology as well as staining behavior after 24 h at 37 and 45°C with the isolated rods some rods were also visible in chains and pairs. Frequency of cells and staining behavior and sporulation of cells and staining behavior and sporulation property was not affected by extremes of pH and at high temperature. At temperature 37°C at pH 6 (~54%) isolated rods, (0.17%) in chains and (0.1%) in pairs were observed, (~45%) spores were also seen. at alkaline pH 9 (37°C) isolated rods were (~39%), (~7%) rods in chains and (~18%) rods in pairs and (~33%) spores were observed. at temperature 37°C rods were decreased from acidic to alkaline pH except at pH 8 (~45%). At temperature 45°C at pH 6 isolated rods (~21%), (~41%)rods in chains and (~18%) rods in pairs were observed. Spores were (~19%). At pH 9 (45%) isolated rods (~36%), (~7%) rods in chains, (~25%) rods in pairs and spores were (~30%). At 45°C the isolated rods were increased with the increase in pH from (6-9), rods in chains were decreased with the increase in pH while rods in pairs (~18-25%) and spores (~19-30%) were increased with the increase in pH in comparison of 45°C with 37°C isolated rods were more at 37°C as compared to 45°C.
| Table 2: | Effect of temperature and pH on bacterial growth of strains PY79, 5, 19, 96 and 99 |
 | |
After 96 h of incubation only gram negative isolated rods and spores were observed at all pHs (6, 7, 8, 9) and at both temperatures (37 and 45°C). At pH 6 and 37°C (~30%) isolated rods and (~70%) spores were observed while at pH 9 with the (~22%) isolated rods, some (~5%) rods in pairs and (~73%) spores were observed.
Strain-SD5
At all pH (6, 7, 8, 9) and at both temperatures (37 and 45°C) all the rods were gram variable. After 24 h at temperature 37°C at pH 6 (~56%) isolated rods, (1.12%) in chains and (~25%) in pairs were observed. (~18%) spores were also seen.
At pH 9 (37°C) isolated rods (~52%), (~12%) rods in pairs and (~64%) spores were observed. After 24 h at temperature 45°C at pH 6 isolated rods (~53%), (~1%) rods in chains and (~27%) rods in pairs were observed. spores were (~19%) but these spores were defective in germination. after 24 h at temperature 45°C at pH 9 isolated rods (~53%), (~17%) rods in pairs and spores were (~30%) present. At 45°C rods in chains were increased with the increase in pH except at pH 9 where they were absent while rods in pairs (~16-27%) and spores were increased with the increase in pH. In comparison of 45°C with 37°C isolated rods and rods in chains were more at 37°C as compared to 45°C. Rods in pairs were more at 45°C then 37°C while spores were more at 37°C as compared to 45°C. After 96 h of incubation at pH 6 and 37°C (~34%) isolated rods, (~11%) rods in pairs and (~55%) spores were observed and at pH 9 (~26%) isolated rods, (~18%) rods in pairs and (~56%) spores were observed. At 45°C at pH 6 (~32%) isolated rods, (~105) rods in pairs and (~58%) spores were observed which were defective while at pH 9 (~20%) isolated rods, (~16%) rods in pairs and (~64%) defective spores were observed.
Strain-SD19
After 24 h of incubation at temperature 37°C and pH 6 (~24%) gram positive isolated rods, (~10%) in chains and (~11%) in pairs were observed. (~55%) spores were also seen. At pH 9 (37°C) isolated rods were (~45%), (~18%) rods in pairs and (~37%) spores were observed. at temperature 37°C rods were more at neutral pH then acidic and alkaline pHs. At temperature 45°C at pH 6 isolated rods (~38%), (~10%) rods in pairs were observed. defective spores (~52%) at above pH and temperature. At pH 9 (45°C) isolated rods (~45%), (~13%) rods in pairs and defective spore were (~42%). At 45°C the isolated rods were more at alkaline pHs then at neutral and acidic pHs while rods in pairs less at extremes of pH and spores were same at all pHs. In comparison of 45°C with 37°C isolated rods were more at 37°C as compared to 45°C while spores were almost same at both temperature (37°C, 45°C) but at 45°C defective spores were observed as compared to 37°C. After 96 h of incubation at pH 6 (37°C), (~25%) isolated rods, (~11%) rods in pairs and (~64%) spores were observed. While at pH 9 with the (~33%) isolated rods, some (~15%) rods in pairs and (~52%) spores were observed.
Strain-SD96
After 24 h of incubation, at temperature 37°C and pH 6 (~27%) isolated rods, (~8%) in pairs were observed. (~65%) ( spores were also observed. At alkaline pH 9 (37°C) isolated rods were (~5%) and (~95%) spores were seen. At temperature 37°C rods were increased from acidic to neutral pH and then to alkaline except at pH 9 (~5%). At temperature 45°C at pH 6 isolated rods (~14%), (~30%) rods in chains and (~16%) rods in pairs were observed. spores were (~40%) which were defective in germination. At pH 9 (45 °C) isolated rods (~27%), (~8%) (rods in chains, (~7%) rods in pairs and defective spores were (~58%). Rods in chains were decreased with the increase in pH while rods in pairs (~6-16%) and spores (~39-7%) increased with the increase in pH. In comparison of 45°C with 37°C isolated rods, rods in chains and rods in pairs were more at 45°C then 37°C while spores were observed more at 45°C. After 96 h of incubation gram negative isolated rods and spores were observed at all pHs (6, 7, 8, 9) and at both temperatures (37 and 45°C). At pH 6 (37°C) (~15%) isolated rods, (~8%) rods in pairs, (~6%) rods in chains and (~71%) spores were observed. While at pH 9 (37°C) (~23%) isolated rods and isolated rods and (~73%) spores were recorded.
Strain-SD99
After 24 h at temperature 37°C and pH 6 (~67%) (isolated rods, (~3%) rods in chains and (~20%) in pairs and (~10%) spores were observed. At alkaline pH 9 (37°C) isolated rods were (~45%), (~12%) rods in chains and (~28%) rods in pairs and spores (~15%) were observed. At temperature 37°C rods were more at acidic and neutral pH while they were less at alkaline pHs. At temperature 45°C at pH 6 isolated rods (~74%), (~14%) rods in pairs were observed. Spores were (~12%) which wee defective. at pH 9 (45°C) isolated rods (27%), (195%) rods in chains, (~29%) rods in pairs and spores were (~25%) which were defective. in comparison of 45°C with 37°C isolated rods were less at 37°C as compared to 45°C. Rods in chain and in pairs were more at 45°C than 37°C while spores were more at 37°C as compared to 45°C. After 96 h of incubation gram negative isolated rods, rods in chains, rods in pairs and spores were observed at all pH (6, 7, 8, 9) and at both temperature (37 and 45°C). At pH 6 (37°C) isolated rods were (~29%), rods in pairs (~5%), rods in chains (~1%) and spores were (~65%) observed. While at pH 9 (~23%) isolated rods, some (~55) rods in pairs, rods in chains (`2%) and (~70%) spores were observed.
DISCUSSION
Bacterial growth rate was monitored at 600 nm after 24 and 96 h of incubation at 37 and 45°C with varying pH (6, 7, 8, 9). After 24 and 96 h of incubation the bacterial growth of strains PY79, 5 (except at pH 7, 37 and 45°C), 19, 96 (except at pH 9, 45°C) and 99 (except at pH 9, 45°C) was more at 37°C (at pH 6, 7, 8, 9) as compared to 45°C (Table 3). Sabri and Hasnain (1994a), Ahmed and Sabri (2004) reported that higher temperature and varying pH affected the growth rate of wild type as well as sporulation defective div mutant strains. The growth of sporulation defective Bacillus strains after 96 h of incubation was less as compared to PY79.
| Table 3: | Frequency of different cell types per 100 cells of bacillus strains(PY79, 5, 19, 96, 99) at different pH (6, 7, 8, 9) at two temperatures (37 and 45°C) after incubating for 24 and 96 h on nutrient broth |
 | |
| * Defective spores | |
The growth rate and sporulation processes are interrelated (Sabri and Hasnain, 1994b). Ahmed and Sabri (2004) also reported that div mutants have low growth rate as compared to PY79. In general growth is affected by extreme of temperature and pH (Bakermans and Nealson, 2004; Richard and Foster, 2004; Yuk and Mashall, 2004; Nascimento et al., 2004). In general at acidic pH the bacterial growth was less as compared to neutral pH. In alkalophiles bacteria the heat treatment at lower pH destroy heat resistance mechanisms of spores and induce low survival rate, optical density decrease and DPA release (Kudo and Horikoshi, 1983). Extreme temperature and pH had adverse effects on cell morphology and cell lysis (Sabri and Hasnain, 1994 a, b). Lysis may be either due to affects of temperatures on biochemical reaction, membrane proteins, membrane lipids and septation (Chaloupka, 1985; Kubitschek, 1990; Intriago and FloodGate, 1991) or due to change in wall components, peptidoglycan and defect in cell envelope (Waximan and Strominger, 1983; Shohayeb and Chopra, 1987).
Strain PY79 after 24 h of incubation shows optimum growth at pH 8 (both at 37 and 45°C). After 24 h of incubation strain 5 showed optimum growth at pH 7 (both at 37 and 45°C). For bacterial strain 19, 96 and 99, pH 7 was favourable for maximal growth at both temperatures (37 and 45°C). pH could also affect the diversity of bacteria (Iqbal, 2000). Cell wall component are important in determining pH tolerance in the medium (Aono et al., 1993).
After 24 h at all pH (6, 7, 8, 9) at both temperatures (37 and 45°C), the maximum number of isolated rods were observed in PY79 (54.92 % at pH 6, 37°C), 5 (56.17 % at pH 6, 37°C), 19 (45.72% at pH 9, 37°C), 96 (51.23 % at pH 8, 45°C) and 99 (74.07 % at pH 6, 45°C). Rods in pairs were more in PY79 (24.01% at pH 8, 45°C), 5 (27.08 % at pH 6, 45°C), 19 (17.14 % at pH 9, 37°C), 96 (16.43% at pH 6, 45°C) and 99 (32.43 % at pH 7, 45°C). Rods in chains were also present in strains PY79 (24.59 % at pH 9, 45°C), 5 (10.25 % at pH 7, 45°C), 19 (9.62 % at pH 6, 37°C), 96 (30.13 % at pH 6, 45°C) and 99 (19.48 % at pH 9, 45°C). Cell growth is influenced by change in temperature (Leroy and Devuyst, 1999). Increase in pH cause variation in shape and size of cell (Lowe et al., 1989). So from above discussion it is clear that cell degeneration and lysis was more in sporulation defective strains (5, 19, 96, 99). Spores, which were formed by the bacterial isolates 5, 19, 96 and 99, were different from wild type. They were unable to germinate.
REFERENCES
- Ahmed, A. and A.N. Sabri, 2004. Effect of varying temperature on growth, morphology and soluble protein content of divI and divII mutant strains of Bacillus subtilis. Sci. Int. (Lahore), 16: 43-52.
Direct Link - Aono, R., M. Ito and K. Horikoshi, 1993. Occurrence of teichuronopeptide in cell walls of group 2 alkaliphilic Bacillus spp. Microbiology, 139: 2739-2744.
CrossRefDirect Link - Bakermans, C. and K.H. Nealson, 2004. Relationship of critical temperature to Macromolecular synthesis and growth yield in Psychrobacter crypegella. J. Bacteriol., 186: 2340-2345.
Direct Link - Butler, Y.X., Y. Abhayawardhane and G.C. Stewart, 1993. Amplification of the Bacillus subtilis maf gene results in arrested septum formation. J. Bacteriol., 175: 3139-3145.
Direct Link - Castanie-Cornet, M.P. and J.W. Foster, 2001. Escherichia coli acid resistance: cAMP receptor protein and a 20 bp cis-acting sequence control pH and stationary phase expression of the gadA and gadBC glutamate decarboxylase genes. Microbiology, 147: 709-715.
Direct Link - Chaloupka, J., 1985. Temperature as a factor regulating the synthesis of microbial enzymes. Microbial. Sci., 2: 86-90.
PubMedDirect Link - Chan, P.F., S.J. Foster, E.T. Ingham and M.O. Clements, 1998. The Staphylococcus aureus alternative sigma factor σB controls the environmental stress response but not starvation survival or pathogenicity in a mouse abscess model. J. Bacteriol., 180: 6082-6089.
Direct Link - Errington, J., R.A. Daniel and D.J. Scheffers, 2003. Cytokinesis in bacteria. Microbiol. Mol. Biol., Rev., 67: 52-65.
Direct Link - Helmann, J.D., M.F.W. Wu, P.A. Kobel, F.J. Gamo and M. Wilson et al., 2001. Global transcriptional response of Bacillus subtilis to heat shock. J. Bacteriol., 183: 7318-7328.
Direct Link - Intriago, P. and G.D, Flood Gate, 1991. Fatty acid composition of estuarine Flexibacter sp. Strain lnp: Effect of salinity, temperature and carbon source for growth. J. Gen. Microbiol., 137: 1503-1509.
Direct Link - Kubitschek, H.E., 1990. Cell growth and abrupt doubling of membrane proteins in Escherichia coli during the division cycle of Escherichia coli. J. Gen. Microbiol., 136: 599-606.
Direct Link - Kuile, B.H., 1994. . Adaptation of the carbon metabolism of Trichomonas vaginalis to the nature and availability of the carbon source. Microbiology, 140: 2503-2510.
Direct Link - Leroy, F. and L. De-Vuyst, 1999. Temperature and pH conditions that prevail during fermentation of sausages are optimal for production of the antilisterial bacteriocin sakacin K. Applied Environ. Microbiol., 65: 974-981.
Direct Link - Nascimento, M.M., J.A.C. Lemos, J. Abranches, R.B. Goncalves and R.A. Burne, 2004. Adaptive acid tolerance response of Streptococcus sobrinus. J. Bacteriol., 186: 6383-6390.
Direct Link - Periago, P.M., 2002. Identification of proteins involved in the heat stress response of Bacillus cereus ATCC 14579. Applied Environ. Microbiol., 68: 3486-3495.
Direct Link - Richard, H. and J.W. Foster, 2004. Escherichia coli glutamate-and arginine-dependant acid resistance system increase internal pH and reserve transmembrane potential. J. Bacteriol., 186: 6032-6041.
Direct Link - Sabri, A.N. and S. Hasnain, 1994. Temperature effects on cell morphology of divIVA+ and divIVA- transformants of B. subtilis. Pak. J. Zool., 26: 149-161.
Direct Link - Schaik, W.V., M.H. Tempelaars, J.A. Wouters, W.M. Devos and T. Abee, 2004. The alternative sigma factor σB of Bacillus cereus: Response to stress and role in heat adaptation. J. Bacteriol., 186: 316-325.
Direct Link - Sadaie, Y., H. Takamatsu, K. Nakamura and K. Yamane, 1991. Sequencing reveals similarity of the wild-type div+ gene of Bacillus subtilis to the Escherichia coli secA gene. Gene, 98: 101-105.
Direct Link - Schut, F., M. Jansen, T.M.P. Gomes, W.H. Gottschal and R.A. Prins, 1995. Substrate uptake and utilization by a marine ultra microbacterium. Microbiology, 141: 351-361.
Direct Link - Stancik, L.M., D.M. Stancik, B. Schimidt, D.M. Barnhart, Y.N. Yoncheva and J.L. Slonzewski, 2002. pH depandant of periplasmic proteins and amino acids metabolism in Escherichia coli. J. Bacteriol., 184: 4246-4258.
Direct Link - Tucker, D.L., N. Tucker, Z. Ma, J.W. Foster, R.L. Miranda, P.S. Cohen and T. Conway, 2003. Genes of the GadX-GadW regulon in Escherichia coli. J. Bacteriol., 185: 3190-3201.
Direct Link - Yanoun, A., R.A. Daniel, J. Errington and C.E. Buchanan, 1993. Cloning and Sequencing of the cell division gene pbpB, which encodes pencillin- binding protein 2B in Bacillus subtilis. J. Bacteriol., 175: 7604-7616.
Direct Link - Yuk, H.G. and D.L. Marshall, 2004. Adaptation of Escherichia coli O157: H 7 to pH alters membrane lipid composition, Verotoxin secretion and resistance to simulated gastric fluid acid. Applied Environ. Microbiol., 70: 3500-3505.
Direct Link