
Pairote Wongputtisin
Biotechnology Program, Graduate School, Chiang Mai University, Thailand
Chartchai Khanongnuch
Department of Biotechnology, Faculty of Agro-Industry, Chiang Mai University, Thailand
Puntipa Pongpiachan
Department of Animal Science, Faculty of Agriculture, Chiang Mai University, Thailand
Saisamorn Lumyong
Department of Biology, Faculty of Science, Chiang Mai University, Thailand
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 7 | Page No.: 577-583







ABSTRACT
The improvement of antioxidant activity of soybean meal (SBM), an important protein source for monogastric animals, was studied by solid state fermentation. Fermented soybean meal (FSBM) was produced using pure culture of each Bacillus subtilis strain MR10 and TK8, B. natto and Rhizopus oligosporus that are accepted as GRAS microorganisms. It was found that the antioxidant activity in term of scavenging effect and reducing power was increased significantly at p<0.05. FSBM produced by Bacillus sp. and R. oligosporus showed percentage scavenging activity at the level of 70-99 and 49%, respectively, while only 25% of scavenging activity was found in SBM. Furthermore, the reducing power value (A700) of every FSBM and SBM was between 0.827-1.031 and 0.299, respectively.
PDF Abstract XML References
How to cite this article
Pairote Wongputtisin, Chartchai Khanongnuch, Puntipa Pongpiachan and Saisamorn Lumyong, 2007. Antioxidant Activity Improvement of Soybean Meal by Microbial Fermentation. Research Journal of Microbiology, 2: 577-583.
URL: https://scialert.net/abstract/?doi=jm.2007.577.583
URL: https://scialert.net/abstract/?doi=jm.2007.577.583
INTRODUCTION
Soybean meal (SBM) is a high quality and the most important source of dietary protein for monogastric animals, especially swine and poultry. It is a by-product obtained after oil extraction process of soybean including drying, dehulling, flaking, expanding, extraction and meal drying. SBM contains 47-51% DM of protein, while the other important nutritional compositions are carbohydrates (35% DM), fat (1.2-3.1% DM) and variety of minerals. However, during oil extraction by solvent, some phytochemicals may be removed from soybean as well, for example vitamin E (α-tocopherol), lecithin and isoflavones. About isoflavovones, there was a report confirmed that it was significantly lost during soaking and heat treatment (Xie and Hettiarachchy, 2001). These compounds function as natural antioxidants preventing many diseases caused by reactive free radicals for human and animals (Lee et al., 2005; Tavva et al., 2006; Yang et al., 2000; Yamamoto et al., 2003). Actually, almost organisms including human and animals are well protected against free radical damage by endogenous enzymes such as superoxide dismutase and catalase, or compounds such as ascorbic acid, tocopherol and glutathione but when this mechanism become imbalance by many factors such as aging or deterioration of physiological functions, pathological stress may occur (Yang et al., 2000). For animals, pathological stress may be found when they are fed in high stress condition or exposed to some chemicals. Supplementation of antioxidants or enhancement of antioxidant in food for human and animals may be used to reduce oxidative stress in body caused by free radicals. Synthetic antioxidants such as butylated hydroxyl anisole (BHA), butylated hydroxyl toluene (BHT), propyl gallate (PG) and tertiary butyl hydroquinone (TBHQ) are widely used because of their effectiveness and cheaper than natural antioxidants. However, their safety and toxicity of synthetic type must be concerned before use. Natural antioxidants are then very interesting source in food and feed application.
Fermented foods represent on average one-third of total food consumption (Nout and Kiers, 2005), especially fermented soybean that are widespread found in many part of the world as a local food for example Tua-nao (Thailand), Natto (Japan), Tempe (Indonesia) and Kinema (India). Those products have been reported on higher in antioxidant activity via microbial fermentation (Nout and Kiers, 2005; Hattori et al., 1995; Yang et al., 2000; Pyo et al., 2005). There are many reports about diversity of microorganisms found in fermented foods, especially in fermented soybean and related products for example Bacillus sp., lactic acid bacteria, Rhizopus oligosporus, Mucor sp., Actinomucor sp. and yeasts (Sarkar et al., 2002; Omafuvbe et al., 2002; Han et al., 2001; Han et al., 2003; Hesseltine, 1998). However, Bacillus sp., especially B. subtilis is the most important producing bacteria found in many fermented soybean products (Hesseltine, 1998). Moreover, R. oligosporus is another microbe that is the main producing mold found in tempe. Both B. subtilis and R. oligosporus have been accepted as GRAS (Generally Regarded as Safe) strain (Fererira et al., 2005; Lin et al., 2006). Not only growing on soybean as sole substrate but those can grow on SBM as well. These microbes were then expected as antioxidant activity enhancer for SBM as the found in fermented soybean. Therefore, in this research, we focused on the improvement of antioxidant activity of SBM for animal feed application using microbial fermentation. Fermented soybean meal (FSBM) production by the selected microorganisms was then studied and the microorganisms used were B. subtilis, B. natto and R. oligosporus isolated from Tua-nao, natto and tempe, respectively. The antioxidant activity in term of scavenging effect and reducing power of SBM and various FSBM will be reported in this study.
MATERIALS AND METHODS
Microorganisms
B. subtilis MR10 and B. subtilis TK8 were isolated from Tua-nao sample in Chiang Mai province, Thailand. They were identified to specie level using the method according to Norris et al. (1981). While B. natto and R. oligosporus were kindly obtained from Microbiology Section, Faculty of Science, Chiang Mai University, Thailand. Bacillus sp. and R. oligosporus were maintained in nutrient agar and potato dextrose agar, respectively. The experiment was conducted at Department of Biology, Chiang Mai University, Thailand during 2006.
FSBM Production and Sample Preparation
Ten gram of SBM was mixed with 20 mL distilled water, mixed well and sterile at 121°C 15 min. After cooling, in case of Bacillus sp., the 105 cfu of 8 h -old starter was transferred to solid medium while for R. oligosporus, the 105 spore was inoculated. After mixing well, these media were incubated at 37°C for 24 h. FSBM were subsequently freeze dried and ground to fine powder for the next step.
For antioxidant extraction, 5 g of powdered sample was mixed well with 70% (v/v) ethanol (Merck®) (prepared by deionized water), filtered through Whatman® paper No. 1. The filtrate was harvested and used as tested crude extract.
Scavenging Effect on ABTS Radical Cation
To generate ABTS•+, the protocol according to Re et al. (1999) was used. The 5 mL 14 mM ABTS (0.0385 g ABTS in 5 mL deionized water) and 5 mL potassium persulfate (0.0066 g potassium persulfate in 5 mL deionized water) were mixed together and stand in the dark for 12-16 h before use. To determine scavenging activity of FSBM extract, 10 μL of extract was added to 990 μL of ABTS•+ solution (adjusted the absorbance at 734 nm to 0.700±0.020 before used) and recorded the decreasing of A734 every 1 min until stable. The standard antioxidants used in this study were α-tocopherol (Merck®), ascorbic acid (Fisher Chemicals®), bultylated hydroxyanisole (BHA, Fluka®). For α-tocopherol and BHA, the solution were prepared in ethanol (Merck®) while ascorbic acid was prepared in 0.2 M sodium phosphate buffer pH 6.5 (Yang et al., 2000). The percent of scavenging activity at 1 min of reaction can be calculated by the formula:
Reducing Power of FSBM Extract
The reducing power of FSBM extracts was determined according to the method of Oyaizu (1986). FSBM extract (2.5 mL) was mixed with sodium phosphate buffer (2.5 mL) and 1% (w/v) potassium ferricyanide (2.5 mL) and subsequently incubated at 50°C 20 min. After that 2.5 mL of 10%(w/v) trichloroacetic acid was added and centrifuged at 3000 rpm 10 min. The 5 mL supernatant was added with 5 mL deionized water and 1 mL of 0.1%(w/v) FeCl2. After exact 2 min of reaction time, the absorbance at 700 nm was measured. A higher in A700 indicates a higher in reducing power.
Statistical Analysis
Data obtained were analyzed statistically by determining standard errors of the mean and analysis of variance using software STATISTIX version 8.
RESULTS AND DISCUSSION
Scavenging Effect on ABTS Radical Cation and Reducing Power
Scavenging effect on ABTS•+ by several crude extract of FSBM and SBM was determined; it was found that all extracts showed scavenging effect on this free radical as shown in Fig. 1. The absorbance at 734 nm was decreased rapidly at the first minute of reaction and slightly decreased until stable. The FSBM extracts showed higher in scavenging rate on ABTS•+. Among these extracts, FSBM produced by B. subtilis TK8 and mixed Bacillus sp. completely removed free radical since 5 and 2 min after reaction time, respectively, while almost complete free radical removal was found from those of B. subtilis MR10 and B. natto.
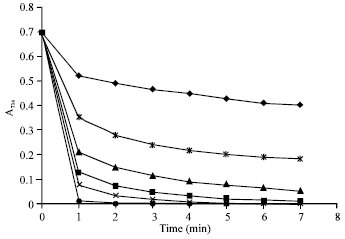 | |
| Fig. 1: | Scavenging effect profile of methanolic extract of SBM and FSBM; SBM (♦), R. oligosporus (*), B. natto (▲), B. subtilis MR10 (■), B. subtilis TK8 (x) and mixed Bacillus sp (●) |
| Table 1: | Scavenging activity of methanolic extract of SBM and FSBMs at 1 min of reaction |
 | |
| Each value represents mean±SD (n = 3) and different letter (s) are significantly different at p<0.05 | |
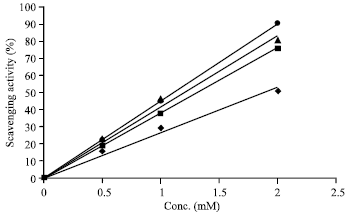 | |
| Fig. 2: | Scavenging effect of standard antioxidants; α-tocopherol (♦), ascorbic acid (●), BHA (▲) and Trolox (■), at various concentration and 1 min of reaction time |
When percentage scavenging activity of the extracts at 1 min of reaction time were calculated, the results showed that FSBM extract of mixed Bacillus sp. and TK8 significantly showed the highest percentage scavenging activity at 98.97 and 91.84%, respectively and the lowest value was found in SBM extract (25.17%) at p<0.05 (Table 1). While FSBM extract of B. subtilis MR10, B. natto and R. oligosporus removed free radical at 82.17, 70 and 49.45%, respectively. Therefore, we found that there was approximately 2-4 times higher in percentage cavenging activity of SBM when microbial fermentation was applied, especially, by the action of Bacillus sp.
The scavenging effect of some standard antioxidants was also investigated to compare with SBM and FSBM extract. It was found that at 1 min of reaction time, FSBM extract of B. subtilis TK8 and mixed Bacillus still exhibited higher in scavenging effect than those of every standard antioxidant at concentration 2 mM. On the other hand, those of SBM extract gave the lower activity than every standard antioxidant. The percentage scavenging activity of α-tocopherol, ascorbic acid, BHA and trolox at 2 mM and 1 min of reaction time were 51.09, 89.75, 80 and 75%, respectively (Fig. 2).
From the results above, we found the fermentation of soybean meal could improve the ability in ABTS•+ removal and we also found the fermentation by bacteria could enhance scavenging activity more satisfy than the one by mold. The difference of growth rate between bacteria and mold might be the reason for this result. Among these Bacillus sp. used in this study, there was a difference in scavenging effect obtained. We found that B. subtilis TK8 gave the best activity when compared to B. subtilis MR10 and B. natto. B. subtilis TK8 and MR10 were isolated from Tua-nao, a traditional fermented soybean of Thailand. B. subtilis TK8 is bacteria that capable of higher in cellulase production than B. subtilis MR10 and B. natto (data and detail not shown). Cell wall of soybean might be hydrolyzed by cellulase activity, therefore better secretion of active compounds from soybean cell was obtained and resulting in better scavenging activity. The active compounds found in soybean seed against free radicals are α-tocopherol, isoflovones and their glycosides such as malonyl glycitin, genistin, daidzin, daidzein, genistein and glycitin (Lee et al., 2005; Pyo et al., 2005; Yamamoto et al., 2003).
| Table 2: | Reducing power of methanolic extract of SBM and FSBMs |
 | |
| Each value represents mean±SD (n = 3) and different letter (s) are significantly different at p<0.005 | |
These active compounds can be found in many parts of soybean seed such as tocopherol that found mainly in cotyledon (92.8%), seed coat (5.4%) and axis (1.8%) (Yoshida et al., 2006), while 80-90% of total isoflavones were located in the cotyledon and the remainder in the hypocotyls (Hoeck et al., 2000). In case of R. oligosporus fermentation, there was a report on increasing of scavenging effect found in tempe caused by this mold (Nout and Kiers, 2005; Lin et al., 2006). That is consistent to our results, although our study was conducted in soybean meal not in whole seeds. The previous reports were studied on the scavenging effect of soybean and related products on several free radicals such as DPPH radical (Yang et al., 2000; Lee et al., 2005; Chen et al., 2005), superoxide anion radical and hydroxyl free radical (Yang et al., 2000; Yen and Hsieh, 1997) but our study showed that soybean meal and FSBMs extract could also eliminate ABTS radical.
Assay of reducing power was based on the reduction of Fe3+/ferricyanide complex to the ferrous form in the presence of reductones (antioxidants) in the tested extracts (Oyaizu, 1986). In this study, the reducing power of the methanolic extracts of SBM and FSBMs was indicated as the higher in the formation of Perl’s Prussian blue (A700) and the results showed that microbial fermentation could improve the reducing power of soybean meal as found in the scavenging effect study. The reducing power of SBM extract was the lowest while the highest reducing power was found in the one of mixed Bacillus sp. at p<0.005. Among FSBM extracts, exception with FSBM extract of mixed Bacillus sp., no significant difference in reducing power of those FSBM extract was found at p<0.005 (Table 2). This result was consistent to their scavenging activity that microbial fermentation enhanced reducing power for SBM as approximately 2-3 times higher was found. In case of R. oligosporus fermentation, we found its reducing power was in the same level as found in extract of koji produced by this mold (Lin et al., 2006). The reducing power of every FSBM might be due to their hydrogen-donating ability as described by Shimada et al. (1992) such as reductone formed during fermentation that could react with active free radicals to stabilize and terminate radical chain reaction (Yang et al., 2000).
On overview of antioxidant activity improvement of SBM using microbial fermentation, many mechanisms might be concerned as same as found in the soybean fermentation. Isoflavones and their glycosides content in SBM and FSBM were considered as a potent scavenging effect and reducing power substances (Lee et al., 2005). Nout and Kiers (2005) reported referring to McCue and Shetty (2003) that during the tempe fermentation, at least a partial cleavage or change in the glucosides take place associated with increased glucosidase and glucuronidase activity, releasing potent antioxidant substances by transformation of flovonoids. Moreover, the other antioxidant substances found in fermented soybean by R. oligosporus were HAA (Esaki et al., 1996) and factor 2 (6, 7, 4-trihydroxyisoflavone) (Nout and Kiers, 2005). Lin et al. (2006) also reported that phenolic compounds that is antioxidants and found in soybean can be increased after fermentation. It was suggested that the liberation of lipophillic aglycones of isoflavone glucosides such as daidzein and genestein by the catalytic action of β-glucosidase during fermentation resulted in the increased antioxidative activity of fermented soybean. Furthermore, there is another mechanism to explain the increasing of antioxidant activity of soybean by fermentation. It might be due to increasing of free amino acid caused by microbial protease activity, especially, tyrosine, methionine, histidine, lysine and tryptophan that are generally accepted to be antioxidants (Wang and Mejia, 2005; Yen and Hsieh, 1997). Wang and Mejia (2005) also reported referring to Chen et al. (2005) that during protein hydrolysis, the soy protein structure will be altered and more active amino acid R group will be exposed. Therefore, soybean peptides can have higher antioxidant activity than intact protein. For example, after enzyme digestion of β-conglycinin and glycinin, the radical-scavenging activity was increased 3-5 times. Those mechanisms given above might be the reason to explain the improvement of SBM by microbial fermentation.
CONCLUSIONS
It could be concluded that the antioxidant activity of SBM, an important protein source for monogastric animals, was improved by the action of tested bacteria and mold that are accepted as GRAS. The scavenging effect on ABTS•+ of every FSBM was approximately 2-4 folds higher than SBM. FSBM fermented by Bacillus sp. exhibited higher scavenging effect than that by Rhizopus oligosporus. For reducing power, the results were in consistent to those of scavenging effect assay that FSBM gave better activity than SBM. In addition, there was no statistically significant difference between FSBM fermented by bacteria and mold. These obtained results suggested that FSBMs might be a potent product for animal feed industry to improve the animal performance.
ACKNOWLEDGMENTS
This research was funded by the Commission on Higher Education, Ministry of Education, Thailand and also supported by a Grant-in-Aid for Graduate student from Graduate School, Chiang Mai University, Chiang Mai, Thailand.
REFERENCES
- Chen, Y.C., Y. Sugiyama, N. Abe, R. Kuruto-Niwa, R. Nozawa and A. Hirota, 2005. DPPH radical-scavenging compounds from dou-chi, a soybean fermented food. Biosci. Biotechnol. Biochem., 69: 999-1006.
Direct Link - Esaki, H., H. Onozaki, S. Kawakishi and T. Osawa, 1996. New antioxidant isolated from tempeh. J. Agric. Food Chem., 44: 696-700.
Direct Link - Fererira, L.C.S., R.C.C. Ferreira and W. Schumann, 2005. Bacillus subtilis as a tool for vaccine development: From antigen factories to delivery vectors. Ann. Acad. Bras. Cienc., 77: 113-124.
Direct Link - Han, B.Z., F.M. Rombouts and M.J.R. Nout, 2001. A Chinese fermented soybean food. Int. J. Food Microbiol., 65: 1-10.
CrossRefDirect Link - Han, B., R.R. Beumer, F.M. Rombouts and M.J.R. Nout, 2001. Microbiological safely and quality of commercial sufu-a Chinese fermented soybean food. Food Control, 12: 541-547.
Direct Link - Hattori, T., H. Ohishi, T. Yokota, H. Ohoami and K. Watanabe, 1995. Antioxidative effect of crude antioxidant preparation from soybean fermented by Bacillus natto. Lebensmittel-Wissenschaft-Technol., 28: 135-138.
Direct Link - Hoeck, J.A., W.R. Fehr, P.A. Murphy and G.A. Welke, 2000. Influence of genotype and environment on isoflavone contents of soybean. Crop Sci., 40: 48-51.
Direct Link - Lee, C.H., L. Yang, J.Z. Xu, S.Y.V. Yeung, Y. Huang and Z.Y. Chen, 2005. Relative antioxidant activity of soybean isoflavones and their glycosides. Food Chem., 90: 735-741.
Direct Link - Lin, C.H., Y.T. Wei and C.C. Chou, 2006. Enhanced antioxidative activity of soybean koji prepared with various filamentous fungi. Food Microbiol., 23: 628-633.
CrossRefDirect Link - McCue, P. and K. Shetty, 2003. Role of carbohydrate-cleaving enzymes in phenolic antioxidant mobilization from whole soybean fermented with Rhizopus oligosporus. Food Biotechnol., 17: 27-37.
CrossRefDirect Link - Nout, M.J.R. and J.L. Kiers, 2005. Tempe fermentation, innovation and functionality: Update into the third millenium. J. Applied Microbiol., 98: 789-805.
CrossRefDirect Link - Omafuvbe, B.O., S.H. Abiose and O.O. Shonukan, 2002. Fermentation of soybean (Glycine max) for soy-daddawa production by starter cultures of Bacillus. Food Microbiol., 19: 561-566.
CrossRefDirect Link - Oyaizu, M., 1986. Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Dietetics, 44: 307-315.
CrossRefDirect Link - Pyo, Y.H., T.C. Lee and Y.C. Lee, 2005. Enrichment of bioactive isoflavones in soymilk fermented with β-glucosidase-producing lactic acid bacteria. Food Res. Int., 38: 551-559.
Direct Link - Re, R., N. Pellegrini, A. Proteggente, A. Pannala, M. Yang and C. Rice-Evans, 1999. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biol. Med., 26: 1231-1237.
CrossRefDirect Link - Sarkar, P.K., B. Hasenack and M.J.R. Nout, 2002. Diversity and functionality of Bacillus and related genera isolated from spontaneously fermented soybeans (Indian Kinema) and locust beans (African Soumbala). Int. J. Food. Microbiol., 77: 175-186.
Direct Link