
M.Y. Elgendy
Department of Hydrobiology, Veterinary Research Division, National Research Centre, Dokki, Giza, Egypt
A.S. Hakim
Department of Microbiology and Immunology, Veterinary Research Division, National Research Centre, Dokki, Giza, Egypt
T.B. Ibrahim
Department of Hydrobiology, Veterinary Research Division, National Research Centre, Dokki, Giza, Egypt
W.S. Soliman
Department of Hydrobiology, Veterinary Research Division, National Research Centre, Dokki, Giza, Egypt
S.E. Ali
Department of Hydrobiology, Veterinary Research Division, National Research Centre, Dokki, Giza, Egypt
Journal of Fisheries and Aquatic Science
Year: 2016 | Volume: 11 | Issue: 3 | Page No.: 206-215







ABSTRACT
The effect of dietary curcumin on the non-specific immune defence mechanisms and resistance against challenge with pathogenic Vibrio alginolyticus in Nile tilapia, Oreochromis niloticus, was evaluated. Fish were divided into two groups before being fed for 30 days with commercial diets supplemented with 0% (control) and 2% curcumin (weight/weight) at the rate of 2% of body weight. Fish were challenged with virulent V. alginolyticus on the 30 days of feeding experiment. Immune parameters including; peroxidase, serum bactericidal activity and serum proteins were investigated to analyze fish immune defence mechanisms. All immune profiles were significantly enhanced in fish fed with curcumin supplemented diets (p<0.05) compared to the control group. Challenge study indicated highest survivability (100%) in the group of fish fed with curcumin whereas, others fed the basal diet showed a survival rate of 40%. The immunomodulatory and antimicrobial effects of curcumin were evaluated also in vitro by assaying Nitric Oxide (NO) production via macrophages and well diffusion method, respectively. A gradual significant increase in nitric oxide production in parallel with higher curcumin dose was demonstrated. The uppermost NO levels, were detected in 50 μg concentration. All curcumin concentrations showed varying degrees of inhibition against tested V. alginolyticus strains and the minimal inhibitory concentration was determined as 12.5 mg mL–1. Therefore, results confirmed substantial evidence to suggest that low concentrations of curcumin as food supplements are able to enhance the immune defence mechanisms of Nile tilapia and could be of value in protection against invading pathogens.
PDF Abstract XML References Citation
Received: November 17, 2015;
Accepted: January 29, 2016;
Published: April 15, 2016
How to cite this article
M.Y. Elgendy, A.S. Hakim, T.B. Ibrahim, W.S. Soliman and S.E. Ali, 2016. Immunomodulatory Effects of Curcumin on Nile Tilapia, Oreochromis niloticus and its Antimicrobial Properties against Vibrio alginolyticus. Journal of Fisheries and Aquatic Science, 11: 206-215.
DOI: 10.3923/jfas.2016.206.215
URL: https://scialert.net/abstract/?doi=jfas.2016.206.215
DOI: 10.3923/jfas.2016.206.215
URL: https://scialert.net/abstract/?doi=jfas.2016.206.215
INTRODUCTION
The global trend of commercial aquaculture is towards intensification of culture practice in the aim to increase productivity per unit area (Elgendy et al., 2015a). Consequently, fish vulnerability to infectious agents is boosted with major stock mortalities and substantial economic losses (Elgendy et al., 2015b, c; Moustafa et al., 2015).
Therefore improvement and innovation of secure methods to combat microbial infections in aquaculture is a critical concern. Traditional approaches include application of chemotherapeutics and antibiotics but the consequential ample undesirable environmental outcomes restrict their therapeutic purposes (Grigorakis and Rigos, 2011). On the other hand, efficient vaccines are not developed against all fish pathogens and their application is quite pricey (Vetvicka et al., 2013).
Strengthening fish defense mechanisms through prophylactic administration of immunostimulants has long been considered a stronghold in the control and management of diseases in aquaculture (Dugenci et al., 2003; Galina et al., 2009). Among the renowned category of such immunostimulants, medicinal herbs, rank first due to their broad spectrum activity, cost effectiveness and environmental-friendly disease preventative measures (Alishahi and Nejad, 2012).
Curcumin is the major biologically active component of turmeric, Curcuma longa and has long been reported to have diverse medicinal properties (Moghadamtousi et al., 2014) including; anti-inflammatory (Bereswill et al., 2010), antioxidant (Al-Jassabi et al., 2012), antimicrobial (Tajbakhsh et al., 2008) and antiviral activities (Kutluay et al., 2008) and immunomodulatory effects (Gupta et al., 2013). Additionally, curcumin has been found to lack significant toxicity in humans even at high oral doses up to 12 g per day (Cheng et al., 2001). Curcumin has been successfully used to control some bacterial infections in farmed fish like Aeromonas hydrophila in goldfish (Harikrishnan and Balasundaram, 2008) and Vibrio harveyi in black tiger shrimp, Penaeus monodon (Vanichkul et al., 2007).
Studies discuss the immunomodulatory effects of dietary curcumin in tilapia fish are scanty. Therefore the present study initiated to investigate such effects on Nile tilapia, Oreochromis niloticus and the potential resistance against challenge with pathogenic V. alginolyticus strain. Further the study aimed to evaluate the immunomodulatory and antimicrobial effects of curcumin in vitro with the determination of the Minimum Inhibitory Concentration (MIC).
MATERIALS AND METHODS
Diet preparation: Curcumin powder was purchased from (Lobal Chemie). A commercial pellet diet was crushed, mixed with tap water containing 2% of curcumin powder (2/100 g of powder) and made again into pellets. Remade pellets were allowed to dry and stored at 4°C.
Fish and experimental design: Nile tilapia, O. niloticus with average weight of 45±5 g were obtained from a private fish farm at Kafer El-Sheikh governorate Egypt. Fish were transported alive to laboratory of Hydrobiology Department, National Research Centre, Egypt, before acclimatized for 15 days in glass aquaria (90×45×45 cm) supplied with chlorine free tap water under continuous aeration. The water was maintained at 25±2°C, pH (7.6), dissolved oxygen (6.6 mg L–1) and ammonia (0.01-0.005 mg L–1). The photoperiod was adjusted to 12 h light:12 h dark. Fish were fed on commercial diet 40% protein at the rate of 2% of body weight.
Fish were divided randomly into 2 groups each with 30 fish (10 per replicate) and fed for 30 days. First group fed with diet supplemented with 2% of curcumin while, other group with basal commercial diets as control.
Determination of immune parameters in vivo
Collection of blood and serum: By the end of feeding experiment, after 30 days, blood samples were obtained from the caudal vein of experimental fish within the two groups for serum analysis and leukocytes isolation. Firstly, fish were anesthetized with clove oil (Merck, Germany) at 50 μL L–1 of water. Blood was drawn via syringe without anticoagulant for serum collection, tubes were kept in slanting position for about 2 h and thereafter centrifuged at 1600xg for 25 min at 4°C, followed by collection of straw colored serum with micropipette and stored at -20°C for further analysis.
Isolation of leukocytes: Leukocytes were isolated from blood according to (Rowley, 1990; Jeney et al., 1997). Briefly, 1 mL of histopaque 1.119 (Sigma, St., Louis, MO) containing bacto hemagglutination buffer, pH 7.3 was dispensed into a siliconized centrifuge tube. Then 1 mL of fish blood was carefully layered on top. The gradient was centrifuged at 500xg for 15 min at 4°C. Interface of leukocyte suspension was gently collected with a Pasteur pipette and dispensed into a siliconized tube before washed twice with phenol red-free Hank’ s balanced salt solution (HBSS, Sigma) and adjusted to 2×106 viable cells per milliliter.
Peroxidase assay: The total peroxidase content in the serum was measured according to Quade and Roth (1997). About 50 μL of different sera obtained from the experimental fish groups was diluted with 135 μL of Hank’s Balanced Salt Solution (HBSS) free from Ca2+ or Mg2+ in 96 well plates. About 25 mL of substrate buffer (Himedia, India) were added. The reaction was stopped after 2 min by adding 50 μL of 4 M sulphuric acid (H2SO4). Plate was c entrifuged at 400xg for 10 min and 150 μL of the supernatants, present in each well, were transferred to new 96 well plates. The OD was recorded at 450 nm in a microplate reader ELx 800 UV (Bio-Tek).
Serum bactericidal activity: Bactericidal activity of sera collected from fish was estimated following the procedure of Kajita et al. (1990). An equal volume (100 μL) of bacterial suspension and serum was mixed and incubated for 1 h at 25°C. Control sample was prepared by replacing fish serum with sterile PBS. The mixture was then diluted with sterile PBS at a ratio 1:10 then 100 μL of the mixture was pour-plated in nutrient agar and incubated for 24 h at 37°C. Bacterial colonies grown in the nutrient agar plates were counted to determine the number of viable bacteria.
Serum proteins: Different sera collected from fish groups were analysed for total protein and albumin content following the methods adopted from Doumas et al. (1971) and Lowry et al. (1951) respectively. Furthermore, globulin content was calculated by subtracting albumin from the total protein then albumin: globulin ratio was determined.
Challenge test: On the 30 days of feeding experiment, each fish within the two groups were intraperitoneally challenged with 0.2 mL of (3×107 CFU) culture suspension of pathogenic V. alginolyticus according to Elgendy (2007). This pathogenic bacterial strain was isolated previously from massive tilapia mortalities by our lab team (Hydrobiology Department National Research Centre, Egypt). Reisolation of injected bacteria from all freshly dead fish during the period of observation was carried out.
The total peroxidase content, serum bactericidal activity and proteins were also determined in sera collected from tilapia fish after the 2 weeks of challenge experiment following the previous described protocols.
In vitro Nitric Oxide (NO) assay: Macrophages were derived from both the head kidneys and peripheral blood according to Soto et al. (2010) and Rajaraman et al. (1998) respectively, then cells were assayed in vitro for Nitric Oxide (NO) production following methods described by Tafalla and Novoa (2000) with some modification. Briefly, after incubation of macrophages the medium was aspirated from the plates. Fifty microliters of Hank's Balanced Salt Solution (HBSS) were added to each well with 50 μL of DMSO in triplicate manner as standard. The two collected macrophage groups were assayed, blood derived macrophages and the head kidney macrophages. Four different curcumin concentrations (150, 50, 15 and 1.5 μg) were added in triplicate manner. The plates were incubated for 2 h at 25°C in a humified 5% CO2 atmosphere. Hundred microliters of the supernatants from each well was transferred into another plate and Hundred microliter of griess reagent were added. The mixtures were incubated at 21°C for 10 min. The Optical density was determined by universal microplate reader ELx 800 UV (Bio-Tek) at 540 NM. The molar concentration of nitrite in the sample was determined from standard curves previously generated with known concentrations of sodium nitrate.
In vitro antimicrobial effects of curcumin against V. alginolyticus: The antibacterial activity of curcumin fraction was screened using the well diffusion method described by Perez et al. (1990). The V. alginolyticus reference strains were obtained from Hydrobiology department, Veterinary division, National Research Centre NRC, Egypt. Colonies of V. alginolyticus were transferred to sterile tubes each containing 5 mL of tryptic soy broth. Turbidity of the bacterial suspensions was adjusted to an optical density equivalent to a 0.5 McFarland standard to give a bacterial suspension of 108 CFU mL–1: Colony Forming Unit.
Mueller-Hinton agar plates were inoculated via streaking bacterial swabs over the entire surface of the plates. Plates were then allowed to dry at room temperature and 6 mm wells were punched in each plate.
Three solutions of known curcumin concentrations were prepared by dissolving exact amount of curcumin fraction into measured volume of ethanol solvent (50, 100 and 400 mg mL–1). Twenty microliters of each concentration were added into wells in each plate. The same amount of solvent, ethanol was also tested.
Plates were then allowed to stand at room temperature to let the tested derivative be diffused into the agar and afterwards, they were incubated at 37°C for 18-24 h, then examined for bacterial growth inhibition and zones of inhibition were measured in millimeters.
Standard streptomycin (30 μg μL–1) and sulphamethoxizole with trimethoprim (1.25/23.75 μg) were used as positive control and sterile water as a negative control for comparison of the antibacterial activity. The experiment was carried out in triplicate and the mean value was recorded.
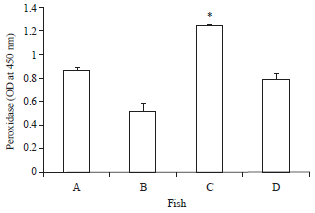 | |
| Fig. 1: | Serum peroxidase activity of O. niloticus fed with curcumin supplemented diet and control basal diet, A: Control fish fed with basal diet, B: Control fish challenged with V. alginolyticus, C: Fish fed with curcumin supplemented diet and D: Fish fed with curcumin supplemented diet after challenge with V. alginolyticus. Data are presented as Mean±SE. Asterisks represent significant difference from control (p<0.05), bars = Mean±SE |
Determination of Minimum Inhibitory Concentration (MIC):
Two-fold serial dilutions were prepared from curcumin solutions in tryptic soy broth. Duplicate tubes of each dilution were inoculated with bacterial strains. All tubes were incubated at 37°C for 18-24 h. The highest dilution of the curcumin that resulted in inhibition of bacterial growth was considered as the MIC.
Statistical analysis: Data were analysed by one-way analysis of variance (ANOVA). The differences among treatments were compared by Tukey's test using Minitab statistical software (Minitab, Coventry, UK). Differences were considered significant at p<0.05.
RESULTS
Effect of curcumin on peroxidase release via O. niloticus macrophages: The peroxidase activity of fish groups varied significantly in the two fish groups (p<0.05). The uppermost peroxidase content was detected in fish fed with curcumin supplemented diet (1.246±0.02) compared to control fish fed only the basal diet (0.860±0.03). Interestingly, after challenge with V. alginolyticus, peroxidase followed a decreasing trend but the curcumin group also showed relatively higher values (0.793±0.05) by comparison to the control group (0.507±0.08) as presented in Fig. 1.
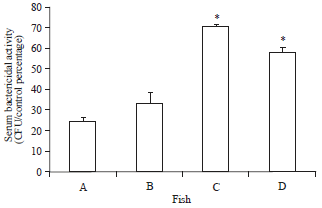 | |
| Fig. 2: | Serum bactericidal activity of O. niloticus fed with curcumin supplemented diet and control basal diet, A: Control fish fed with basal diet, B: Control fish challenged with V. alginolyticus, C: Fish fed with curcumin supplemented diet and D: Fish fed with curcumin supplemented diet after challenge with V. alginolyticus. Data are presented as Mean±SE. Asterisks represent significant difference from control (p<0.05), bars = Mean±SE |
Serum bactericidal activity: The serum bactericidal activity was significantly increased in tilapia fed with curcumin supplemented diet compared to control group including the post challenge period. The differences were significant (p<0.05) as compared with control. The highest amount of serum bactericidal activity was recorded on 30th day in the groups fed curcumin 70.41%±1.30 (CFU/control percentage) compared to control 24.83%±1.97 (CFU/control percentage). Bactericidal activity followed also an increasing trend in curcumin group 58.08%±2.38 after challenge with V. alginolyticus in comparison with fish fed with the basal diet 33.08%±5.71 (Fig. 2).
Serum proteins: The total protein, albumen and globulin increased significantly with the administration of curcumin in comparison with the control group including the post challenge period. On the contrary, the Albumin/Globulin (A/G) ratio was the lowest in the group of fish fed with curcumin (0.49±0.01) in comparison to others fed the basal diet (1.52±0.12) (Fig. 3-6).
Protection upon challenge: The survivability was found highest (100%) in the group fed with curcumin whereas the control group showed a survival rate of 40%. Furthermore, V. alginolyticus was re-isolation from all dead fish.
Nitric Oxide (NO) assay: The macrophage NO production was significantly enhanced after in vitro incubation of macrophages with different curcumin concentrations; 150, 50, 15 and 1.5 μg.
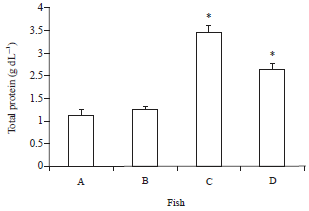 | |
| Fig. 3: | Total serum protein of O. niloticus fed with curcumin supplemented diet and control basal diet A: Control Fish fed with basal diet, B: Control fish challenged with V. alginolyticus, C: Fish fed with curcumin supplemented diet and D: Fish fed with curcumin supplemented diet after challenge with V. alginolyticus. Data are presented as Mean±SE. Asterisks represent significant difference from control (p<0.05), bars = Mean±SE |
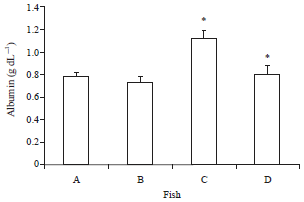 | |
| Fig. 4: | Serum albumen of O. niloticus fed with curcumin supplemented diet and control basal diet, A: Control fish fed with basal diet, B: Control fish challenged with V. alginolyticus, C: Fish fed with curcumin supplemented diet and D: Fish fed with curcumin supplemented diet after challenge with V. alginolyticus. Data are presented as Mean±SE. Asterisks represent significant difference from control (p<0.05), bars = Mean±SE |
The NO production followed a gradual significant increasing trend parallel with higher crucumin concentration. The uppermost generation of NO was observed in 50 μg crucumin while, 150 μg achieved a non significant increase.
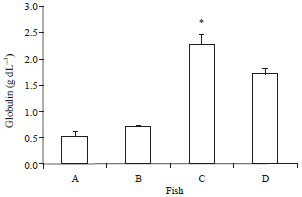 | |
| Fig. 5: | Serum globulin of O. niloticus fed with curcumin supplemented diet and control basal diet, A: Control fish fed with basal diet, B: Control fish challenged with V. alginolyticus, C: Fish fed with curcumin supplemented diet and D: Fish fed with curcumin supplemented diet after challenge with V. alginolyticus. Data are presented as Mean±SE. Asterisks represent significant difference from control (p<0.05), bars = Mean±SE |
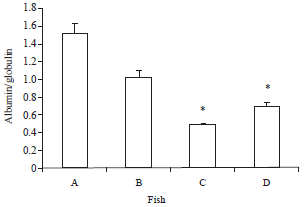 | |
| Fig. 6: | Albumen/globulin ratio of O. niloticus fed with curcumin supplemented diet and control basal diet, A: Control fish fed with basal diet, B: Control fish challenged with V. alginolyticus, C: Fish fed with curcumin supplemented diet and D: Fish fed with curcumin supplemented diet after challenge with V. alginolyticus. Data are presented as Mean±SE. Asterisks represent significant difference from control (p<0.05), bars = Mean±SE |
Moreover, head kidneys derived macrophages demonstrated the highest values of NO production in comparison with peripheral blood macrophages (Fig. 7 and 8).
In vitro antimicrobial effects and minimum inhibition concentration: All prepared curcumin concentrations (50, 100 and 400 mg mL–1) showed varying degrees of inhibition against tested V. alginolyticus strains with zone of inhibition ranged as 13, 15 and 19 mm, respectively.
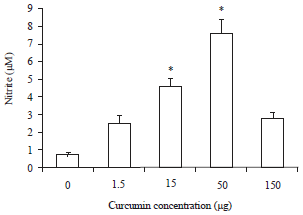 | |
| Fig. 7: | Effect of different curcumin concentrations on nitric oxide production via head-kidney macrophages. Data are presented as Mean±SE. Asterisks represent significant difference from control (p<0.05), bars = Mean±SE |
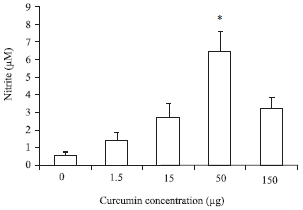 | |
| Fig. 8: | Effect of different curcumin concentration on nitric oxide production via peripheral blood macrophages. Data are presented as Mean±SE. Asterisks represent significant difference from control (p<0.05), bars = Mean±SE |
The MIC was found to be ranging between 12.5 mg mL–1. The results of the two reference antibiotics streptomycin and sulphamethoxizole/trimethoprim compound also showed activity against V. alginolyticus with inhibition zone of about 13 and 15 mm respectively on the other hand absolute ethanol and water were inactive (Fig. 9).
DISCUSSION
Interest in the application of immunostimulants as an alternative method to chemicals and antimicrobials currently being utilized to control fish diseases is growing. The use of medicinal plant products as potential therapeutic measures for modulating the immune mechanisms as well as to control fish diseases have been focused on (Chakrabarti et al., 2014).
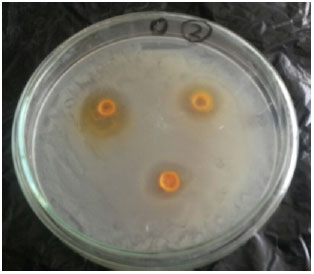 | |
| Fig. 9: | In vitro antimicrobial effects of curcumin against V. alginolyticus |
Numerous herbs are presently used in commercial aquaculture as potent antimicrobial agents, nutrients as well as growth-promoting substances (Vaseeharan and Thaya, 2014; Yilmaz and Ergun, 2014).
Results demonstrated that peroxidase content was increased significantly in the group of fish fed with curcumin supplemented diet in comparison with control fish fed basal diet (p<0.05). The peroxidase produced by fish macrophages are alleged to play significant roles in innate immune-defenses mechanisms against various fish pathogens (Stosik et al., 2001) as well as has been considered as a reliable measure for neutrophilic antimicrobial activity particularly the primary granule exocytosis (Quade and Roth, 1997).
In the presence of halide and hydrogen peroxide, peroxidase generates oxidants destroy pathogens by halogenation of their cell wall (Fischer et al., 2006). The hypochlorous acid produced by peroxidase is a potent bactericidal (Klebanoff, 1967) and markedly potentiates the antibacterial potency of other reactive oxygen species involved in fish defenses (Rosen et al., 1990). Inadequate peroxidase activity in fish fed with the basal diet may indicate poor host defense mechanisms. In agreement with this study, enhancement of peroxidase activity in fish fed with different immunostimulant supplements have been reported likewise; Achyranthes aspera fed to Labeo rohita, Cyprinus carpio (Chakrabarti et al., 2012) and andrographolide in Labeo rohita (Basha et al., 2013).
The serum bactericidal activity was significantly increased in fish group fed with curcumin supplemented diet comparedto the control group including the post challenge period (p<0.05). The bactericidal activity has been viewed as an important tool to analyze the innate immune system. It evaluates the presence of protective proteins in fish blood. Augmented serum bactericidal activities indicate elevated humoral factors involved in the innate and/or adaptive immune mechanisms (Wang et al., 2010). Diverse immunostimulants have been found to promote the serum bactericidal activity. Epinphelus tauvina, fed diets supplemented with extract mixture of some herbs; Cynodon dactylon, Piper longum, Phyllanthus niruri, Tridax procumbens and Zingiber officinalis showed a significant increase in their serum bactericidal activity (Punitha et al., 2008). Additionally, dietary administration of Zingiber officinale enhanced Indian major carp, Catla catla, serum bactericidal activities (Arulvasu et al., 2013).
The present results revealed significant increase in total protein, albumen and globulin levels in Nile tilapia fed with curcumin supplemented diet compared to control fish including the post challenge period (p<0.05). Similar results were detected in Indian major carp, Catla catla, on dietary administration of Zingiber officinale (Arulvasu et al., 2013) as well as in Labeo rohita fed with Magnifera indica kernel (Sahu et al., 2007).
Serum proteins are considered a fundamental index of fish health. Enhanced serum total proteins and globulin levels are thought to associate with stronger fish innate immune response mechanisms (Wiegertjes et al., 1996). Globulins in particular, play a significant role in the immune-protective mechanisms of fish (Sahoo and Mukherjee, 2001) and constitute integral part of almost all the immunoglobulins of blood (Misra et al., 2006).
The Albumin/Globulin (A/G) ratio has long been considered a reliable measure for the humoral components of nonspecific immune defenses (Misra et al., 2006). The low A/G ratio in curcumin treated group reflects overproduction of globulins, with significance for enhanced immune-protective mechanisms induced by the stimulatory effect of curcumin on fish non specific immune responses. On the other hand the high A/G ratio noticed in fish fed with the basal diet suggests underproduction of immunoglobulins. Similarly, feeding Labeo rohita fish with levan supplemented diets enhanced the non-specific cell mediated immune parameters and significantly reduced the A/G ratio (Gupta et al., 2008).
Survivability following challenge with V. alginolyticus was highest (100%) in fish fed with curcumin, whereas the control group showed a survival rate of 40%. Similarly, Nile tilapia fed a mixture of Astragalus and Lonicera extracts had an enhanced immune defence mechanisms as well as increased survival against challenge with Aeromonas hydrophila (Ardo et al., 2008). Moreover, intraperitoneal injection of Oreochromis mossambicus with water soluble fraction of Solanum trilobatum significantly enhanced production of reactive oxygen species and decreased fish mortality following a challenge with A. hydrophila (Divyagnaneswari et al., 2007).
Using fish primary macrophages as a model to investigate the dynamic response process of immunological parameters is highly valued. In vitro experiment demonstrated that NO production was significantly enhanced after incubation of macrophages exposed to heat killed V. alginolyticus with different curcumin concentrations; 150, 50, 15 and 1.5 μg. NO production followed a gradual significant increasing trend parallel with higher crucumin concentration. The uppermost synthesize of NO was observed in 50 μg crucumin while, 150 μg achieved a non significant increase. Moreover, head kidneys derived macrophages demonstrated the highest values of NO production in comparison with blood macrophages. Esteban and Meseguer (1997) have reported that defensive mechanisms of fish macrophages are functionally influenced by tissue source and variation is relevant to their maturation stage. Additionally, studies also demonstrated that monocytes freshly isolated from blood need a long to functionally mature (Shaala et al., 1979).
Diverse immune stimulating substances have been found to modulate the functions of macrophages in vitro. Exposure of rainbow trout, Oncorhynchus mykiss, macrophages retrieved from head kidneys and spleen with the immunomodulator muramyl dipeptide (MDP) in vitro resulted in a significant time-dependent accumulation of NO in cultures (Zvizdic et al., 2012).
Nitric oxides do many crucial roles in the immune defence mechanisms as well as having a direct antimicrobial effect (Villamil et al., 2002). The NOs are considered as potent free oxygen radicals and strongly involved in the pathways of a broad spectrum of diseases performing as a cytotoxic agent in pathological processes (Bogdan et al., 2000; Bogdan, 2000; Aktan, 2004). Furthermore, NOs can inhibit diversity of pathogens including, viruses, bacteria, parasites and many fungal infections (Yang et al., 2013). Additionally, NOs mediate the ability of macrophages to kill or inhibit the growth of many pathogens since it accumulates inside phagocytic vacuoles until it reaches the concentration required to produce the bactericidal effect (Bogdan, 2001).
Regarding the in vitro antimicrobial effects of curcumin against V. alginolyticus, prepared curcumin concentrations showed varying degrees o f inhibition against tested V. alginolyticus strains and 12.5 mg mL–1 was identified as the MIC. The antimicrobial activities of curcumin and turmeric extracts against different bacteria have been reported in some previous studies (Moghadamtousi et al., 2014). The C. longa was found to have inhibitory effect against some bacterial strains including; A. hydrophila (Harikrishnan and Balasundaram, 2008), Helicobacter pylori (Zaidi et al., 2009), Pseudomonas aeruginosa (Negi et al., 1999), E. coli (Gupta and Ravishankar, 2005), Listeria monocytogenes, Salmonella typhimurium and methicillin-resistant Staphylococcus aureus (Kim et al., 2005) as well as K. pneumoniae and Staphylococcus epidermidis (Niamsa and Sittiwet, 2009). The mechanisms of curcumin antimicrobial effects are relevant to suppression of bacterial cell proliferation (Rai et al., 2008) as well as disruption of prokaryotic cell division (Kaur et al., 2010).
CONCLUSION
Fish survival in the aquatic environment requires a competent immune system to overcome the constant challenge with pathogens. Results showed that curcumin can be a promising candidate for immunostimulation as well as a rival substitute for many antimicrobials currently used in tilapia fish farming. The activation of non-specific immune profiles in fish fed with curcumin supplemented diet has direct evidence o n improved health status of the host that might have maintained protection against challenge with V. alginolyticus. Prophylactic administration of dietary curcumin could be of value in protection against invading pathogens as well as minimizing mortalities stemming from disease outbreaks attacking farmed aquatic species.
REFERENCES
- Aktan, F., 2004. iNOS-mediated nitric oxide production and its regulation. Life Sci., 75: 639-653.
CrossRefDirect Link - Alishahi, M. and A.H.J. Nejad, 2012. Effects of propolis, a honeybee product, on growth performance and immune responses of Barbus barbulus. Iran. J. Vet. Med., 6: 249-257.
Direct Link - Al-Jassabi, S., K.A.A. Ahmed and M.A. Abdulla, 2012. Antioxidant effect of curcumin against microcystin-LR-induced renal oxidative damage in Balb/c mice. Trop. J. Pharmaceut. Res., 11: 531-536.
Direct Link - Ardo, L., G. Yin, P. Xu, L. Varadi, G. Szigeti, Z. Jeney and G. Jeney, 2008. Chinese herbs (Astragalus membranaceus and Lonicera japonica) and boron enhance the non-specific immune response of Nile tilapia (Oreochromis niloticus) and resistance against Aeromonas hydrophila. Aquaculture, 275: 26-33.
CrossRefDirect Link - Arulvasu, C., K. Mani, D. Chandhirasekar, D. Prabhu and S. Sivagnanam, 2013. Effect of dietary administration of Zingiber officinale on growth, survival and immune response of Indian major carp, Catla catla (Ham.). Int. Pharm. Pharmaceut. Sci., 5: 108-115.
Direct Link - Basha, K.A., R.P. Raman, K.P. Prasad, K. Kumar, E. Nilavan and S. Kumar, 2013. Effect of dietary supplemented andrographolide on growth, non-specific immune parameters and resistance against Aeromonas hydrophila in Labeo rohita (Hamilton). Fish Shellfish Immunol., 35: 1433-1441.
CrossRefDirect Link - Bereswill, S., M. Munoz, A. Fischer, R. Plickert and L.M. Haag et al., 2010. Anti-inflammatory effects of resveratrol, curcumin and simvastatin in acute small intestinal inflammation. PLoS ONE, Vol. 5.
CrossRefDirect Link - Bogdan, C., 2000. The Function of Nitric Oxide in the Immune System. In: Nitric Oxide (Handbook of Experimental Pharmacology, Volume 143), Mayer, B. (Ed.). Chapter 18, Springer, Berlin, Germany, ISBN: 978-3-642-63026-2, pp: 443-492.
Direct Link - Bogdan, C., M. Rollinghoff and A. Diefenbach, 2000. The role of nitric oxide in innate immunity. Immunol. Rev., 173: 17-26.
CrossRefDirect Link - Bogdan, C., 2001. Nitric oxide and the immune response. Nat. Immunol., 2: 907-916.
CrossRefDirect Link - Chakrabarti, R., P.K. Srivastava, K. Kundu, R.S. Khare and S. Banerjee, 2012. Evaluation of immunostimulatory and growth promoting effect of seed fractions of Achyranthes aspera in common carp Cyprinus carpio and identification of active constituents. Fish Shellfish Immunol., 32: 839-843.
CrossRefDirect Link - Chakrabarti, R., P.K. Srivastava, N. Verma and J. Sharma, 2014. Effect of seeds of Achyranthes aspera on the immune responses and expression of some immune-related genes in carp Catla catla. Fish Shellfish Immunol., 41: 64-69.
CrossRefDirect Link - Cheng, A.L., C.H. Hsu, J.K. Lin, M.M. Hsu and Y.F. Ho et al., 2001. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res., 21: 2895-2900.
PubMedDirect Link - Divyagnaneswari, M., D. Christybapita and R.D. Michael, 2007. Enhancement of nonspecific immunity and disease resistance in oreochromis mossambicus by Solanum trilobatum leaf fractions. Fish Shellfish Immunol., 23: 249-259.
CrossRefDirect Link - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - Dugenci, S.K., N. Arda and A. Candan, 2003. Some medicinal plants as immunostimulant for fish. J. Ethnopharmacol., 88: 99-106.
CrossRefPubMedDirect Link - Elgendy, M.Y., M. Abdelsalam, M. Moustafa, A.M. Kenawy and A. Seida, 2015. Caligus elongatus and Photobacterium damselae subsp piscicida concomitant infections affecting broodstock European seabass, Dicentrarchus labrax, with special reference to histopathological responses. J. Aquacult. Res. Dev., Vol. 6.
CrossRefDirect Link - Elgendy, M.Y., M. Moustafa, A.Y. Gaafar and T.B. Ibrahim, 2015. Impacts of extreme cold water conditions and some bacterial infections on earthen-pond cultured Nile tilapia, Oreochromis niloticus. Res. J. Pharm. Biol. Chem. Sci., 6: 136-145.
Direct Link - Elgendy, M.Y., W.S. Soliman, H.A. Hassan, A.M. Kenawy and A.M. Liala, 2015. Effect of abrupt environmental deterioration on the eruption of vibriosis in mari-cultured shrimp, Penaeus indicus, in Egypt. J. Fish. Aquatic Sci., 10: 146-158.
Direct Link - Esteban, M.A. and J. Meseguer, 1997. Factors influencing phagocytic response of macrophages from the sea bass (Dicentrarchus labrax L.): An ultrastructural and quantitative study. Anat. Rec., 248: 533-541.
PubMedDirect Link - Fischer, U., K. Utke, T. Somamoto, B. Kollner, M. Ototake and T. Nakanishi, 2006. Cytotoxic activities of fish leucocytes. Fish Shellfish Immunol., 20: 209-226.
CrossRefDirect Link - Galina, J., G. Yin, L. Ardo and Z. Jeney, 2009. The use of immunostimulating herbs in fish: An overview of research. Fish Physiol. Biochem., 35: 669-676.
CrossRefDirect Link - Grigorakis, K. and G. Rigos, 2011. Aquaculture effects on environmental and public welfare-The case of Mediterranean mariculture. Chemosphere, 85: 899-919.
CrossRefDirect Link - Gupta, S. and S. Ravishankar, 2005. A comparison of the antimicrobial activity of garlic, ginger, carrot and turmeric pastes against Escherichia coli O157: H7 in laboratory buffer and ground beef. Foodborne. Pathog. Dis., 2: 330-340.
CrossRefPubMedDirect Link - Gupta, S.C., S. Patchva and B.B. Aggarwal, 2013. Therapeutic roles of curcumin: Lessons learned from clinical trials. Am. Associ. Pharm. Sci. J., 15: 195-218.
CrossRefDirect Link - Gupta, S.K., A.K. Pal, N.P. Sahu, R. Dalvi, V. Kumar and S.C. Mukherjee, 2008. Microbial levan in the diet of Labeo rohita Hamilton juveniles: Effect on non-specific immunity and histopathological changes after challenge with Aeromonas hydrophila. J. Fish Dis., 31: 649-657.
CrossRefDirect Link - Harikrishnan, R. and C. Balasundaram, 2008. In vitro and in vivo studies of the use of some medicinal herbals against the pathogen Aeromonas hydrophila in goldfish. J. Aquat. Anim. Health, 20: 165-176.
CrossRefPubMedDirect Link - Jeney, G., M. Galeotti, D. Volpatti, Z. Jeney and D.P. Anderson, 1997. Prevention of stress in rainbow trout (Oncorhynchus mykiss) fed diets containing different doses of glucan. Aquaculture, 154: 1-15.
CrossRefDirect Link - Kajita, Y., M. Sakai, S. Atsuta and M. Kobayash, 1990. The immunomodulatory effects of levamisole on rainbow trout, Oncorhynchus mykiss. Fish Pathol., 25: 93-98.
Direct Link - Kaur, S., N.H. Modi, D. Panda and N. Roy, 2010. Probing the binding site of curcumin in Escherichia coli and Bacillus subtilis FtsZ-A structural insight to unveil antibacterial activity of curcumin. Eur. J. Med. Chem., 45: 4209-4214.
CrossRefDirect Link - Kim, K.J., H.H. Yu, J.D. Cha, S.J. Seo, N.Y. Choi and Y.O. You, 2005. Antibacterial activity of Curcuma longa L. against methicillin-resistant Staphylococcus aureus. Phytother. Res., 19: 599-604.
CrossRefPubMedDirect Link - Klebanoff, S.J., 1967. Iodination of bacteria: A bactericidal mechanism. J. Exp. Med., 126: 1063-1078.
CrossRefPubMedDirect Link - Kutluay, S.B., J. Doroghazi, M.E. Roemer and S.J. Triezenberg, 2008. Curcumin inhibits herpes simplex virus immediate-early gene expression by a mechanism independent of p300/CBP histone acetyltransferase activity. Virology, 373: 239-247.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Misra, C.K., B.K. Das, S.C. Mukherjee and P. Pattnaik, 2006. Effect of long term administration of dietary β-glucan on immunity, growth and survival of Labeo rohita fingerlings. Aquaculture, 255: 82-94.
CrossRefDirect Link - Moghadamtousi, S.Z., H. Abdul Kadir, P. Hassandarvish, H. Tajik, S. Abubakar and K. Zandi, 2014. A review on antibacterial, antiviral and antifungal activity of curcumin. BioMed Res. Int., Vol. 2014.
CrossRefDirect Link - Moustafa, M., A.E. Eissa, A.M. Laila, A.Y. Gaafar, I.M.K. Abumourad and M.Y. Elgendy, 2015. Investigations into the potential causes of mass kills in mari-cultured Gilthead sea bream, (Sparus aurata) at Northern Egypt. Res. J. Pharm. Biol. Chem. Sci., 6: 466-477.
Direct Link - Negi, P.S., G.K. Jayaprakasha, L.J.M. Rao and K.K. Sakariah, 1999. Antibacterial activity of turmeric oil: A byproduct from curcumin manufacture. J. Agric. Food. Chem., 47: 4297-4300.
CrossRefDirect Link - Niamsa, N. and C. Sittiwet, 2009. Antimicrobial activity of Curcuma longa aqueous extract. J. Pharmacol. Toxicol., 4: 173-177.
CrossRefDirect Link - Perez, C., M. Pauli and P. Bazerque, 1990. An antibiotic assay by the agar-well diffusion method. Acta Biol. Med. Exp., 15: 113-115.
Direct Link - Punitha, S.M.J., M.M. Babu, V. Sivaram, V.S. Shankar and S.A. Dhas et al., 2008. Immunostimulating influence of herbal biomedicines on nonspecific immunity in grouper Epinephelus tauvina juvenile against Vibrio harveyi infection. Aquacult. Int., 16: 511-523.
CrossRefDirect Link - Quade, M.J. and J.A. Roth, 1997. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol., 58: 239-248.
CrossRefPubMedDirect Link - Rai, D., J.K. Singh, N. Ray and D. Panda, 2008. Curcumin inhibits FtsZ assembly: An attractive mechanism for its antibacterial activity. Biochem. J., 410: 147-155.
CrossRefPubMedDirect Link - Rajaraman, V., B.J. Nonnecke, S.T. Franklin, D.C. Hammell and R.L. Horst, 1998. Effect of vitamins A and E on nitric oxide production by blood mononuclear leukocytes from neonatal calves fed milk replacer. J. Dairy Sci., 81: 3278-3285.
CrossRefDirect Link - Rosen, H., J. Orman, R.M. Rakita, B.R. Michel and D.R. VanDevanter, 1990. Loss of DNA-membrane interactions and cessation of DNA synthesis in myeloperoxidase-treated Escherichia coli. Proc. Natl. Acad. Sci. USA., 87: 10048-10052.
PubMedDirect Link - Sahoo, P.K. and S.C. Mukherjee, 2001. Effect of dietary β-1,3 glucan on immune responses and disease resistance of healthy and aflatoxin B1-induced immunocompromised rohu (Labeo rohita Hamilton). Fish Shellfish Immunol., 11: 683-695.
CrossRefDirect Link - Sahu, S., B.K. Das, B.K. Mishra, J. Pradhan and N. Sarangi, 2007. Effect of Allium sativum on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila. J. Applied Ichthyol., 23: 80-86.
CrossRefDirect Link - Shaala, A.Y., H.S. Dhaliwal, S. Bishop and N.R. Ling, 1979. Ingestion of dyed-opsonised yeasts as a simple way of detecting phagocytes in lymphocyte preparations. Cytophilic binding of immunoglobulins by ingesting cells. J. Immunol. Methods, 27: 175-187.
CrossRefDirect Link - Soto, E., D. Fernandez, R. Thune and J.P. Hawke, 2010. Interaction of Francisella asiatica with Tilapia (Oreochromis niloticus) innate immunity. Infect. Immunity, 78: 2070-2078.
CrossRefDirect Link - Stosik, M., W. Deptula and M. Travnicek, 2001. Resistance in carps (Cyprinus Carpio L.) affected by a natural bacterial infection. Veterinarni Medicina, 46: 6-11.
Direct Link - Tafalla, C. and B. Novoa, 2000. Requirements for nitric oxide production by turbot (Scophthalmus maximus) head kidney macrophages. Dev. Comparat. Immunol., 24: 623-631.
CrossRefDirect Link - Tajbakhsh, S., K. Mohammadi, I. Deilami, K. Zandi, M. Fouladvand, E. Ramedani and G. Asayesh, 2008. Antibacterial activity of indium curcumin and indium diacetylcurcumin. Afr. J. Biotechnol., 7: 3832-3835.
Direct Link - Vanichkul, K., N. Areechon and N. Kongkathip, 2007. Application of turmeric (Curcuma longa Linn.) extract in black tiger shrimp (Penaeus monodon Fabricius) culture. Proceedings of the 45th Kasetsart University Annual Conference on Self-Sufficient Agriculture: Projects Initiated by His Majesty the King, January 30-February 2, 2007, Kasetsart University, Bangkok, pp: 212-220.
- Vaseeharan, B. and R. Thaya, 2014. Medicinal plant derivatives as immunostimulants: an alternative to chemotherapeutics and antibiotics in aquaculture. Aquacult. Int., 22: 1079-1091.
CrossRefDirect Link - Vetvicka, V., L. Vannucci and P. Sima, 2013. The effects of β-glucan on fish immunity. North Am. J. Med. Sci., 5: 580-588.
CrossRefPubMedDirect Link - Villamil, L., C. Tafalla, A. Figueras and B. Novoa, 2002. Evaluation of immunomodulatory effects of lactic acid bacteria in turbot (Scophthalmus maximus). Clin. Diagn. Lab. Immunol., 9: 1318-1323.
CrossRefPubMedDirect Link - Wang, G.X., Y.T. Liu, F.Y. Li, H.T. Gao, Y. Lei and X.T. Liu, 2010. Immunostimulatory activities of bacillus simplex dr-834 to carp (Cyprinus carpio). Fish Shellfish Immunol., 29: 378-387.
CrossRefPubMedDirect Link - Wiegertjes, G.F., R.J.M. Stet, H.K. Parmentier and W.B. van Muiswinkel, 1996. Immunogenetics of disease resistance in fish: A comparative approach. Dev. Comparat. Immunol., 20: 365-381.
CrossRefDirect Link - Yang, J., R. Zhang, G. Lu, Y. Shen and L. Peng et al., 2013. T cell-derived inducible nitric oxide synthase switches off TH17 cell differentiation. J. Exp. Med., 210: 1447-1462.
CrossRefDirect Link - Yilmaz, S. and S. Ergun, 2014. Dietary supplementation with allspice Pimenta dioica reduces the occurrence of streptococcal disease during first feeding of mozambique tilapia fry. J. Aquat. Anim. Health, 26: 144-148.
CrossRefDirect Link - Zaidi, S.F., K. Yamada, M. Kadowaki, K. Usmanghani and T. Sugiyama, 2009. Bactericidal activity of medicinal plants, employed for the treatment of gastrointestinal ailments, against Helicobacter pylori. J. Ethnophamacol., 121: 286-291.
CrossRefPubMedDirect Link - Zvizdic, M.Z., E. Licek and C. Lam, 2012. Muramyl dipeptide, a stimulator of nonspecific immunity, induces the production of nitric oxide by Rainbow trout (Oncorhynchus mykiss) spleen and head kidney cells in vitro. Vet. Arhiv., 82: 617-627.
Direct Link