
A. Norashikin
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Z. Muta Harah
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
B. Japar Sidik
Faculty of Agriculture and Food Sciences, Universiti Putra Malaysia Bintulu Sarawak Campus, 97008 Bintulu, Sarawak, Malaysia
Journal of Fisheries and Aquatic Science
Year: 2013 | Volume: 8 | Issue: 3 | Page No.: 452-461







ABSTRACT
Intertidal areas exhibit wide ranges in their extent and environmental conditions and these potentially can result in complex distributional pattern and life styles for many species including seaweeds. A study on seaweeds diversity and life forms was conducted at two intertidal sites in Negeri Sembilan i.e., (1) Teluk Kemang, a rocky outcrops area extending up to 50 m seaward during low tide, with gradual slope with live and dead corals interspersed with sand and (2) Teluk Pelanduk, a stretch of up to 150 m flat beach with mostly mud, sand and little rocks in between. A total of 36 species (16 Chlorophyta, 10 Phaeophyta and 10 Rhodophyta) and 26 species (11 Chlorophyta, 6 Phaeophyta and 9 Rhodophyta) of seaweeds were identified at Teluk Kemang and Teluk Pelanduk, respectively. Jaccard similarity coefficient of 0.442 based on a presence-absence matrix suggests the seaweed composition and diversity varied between the two sites. Both sites possessed their own many specialist species. Variability in environmental conditions e.g., diverse natural substrates or abandoned drift objects (fishing nets and lines, ropes and tyres) introduced by human had caused variation in seaweed composition, diversity and life forms categories. Epilithic (45.71%) seaweed is the most abundant category at Teluk Kemang followed by, epizoic (24.29%), epipelic (12.86%), epiphytic (8.57%) and drift seaweed (8.57%). Seaweed life forms at Teluk Pelanduk are epilithic (38.46%), epipelic (21.15%), epiphytic (17.31%), drift seaweed (13.46%) and epizoic (9.62%). Seaweed species distribution irrespective of sites reflected those that adapt or have adapted to the environmental conditions, particularly in utilizing the availability of the diverse existing natural hard and soft substrates and abandoned drift objects.
PDF Abstract XML References Citation
Received: September 19, 2012;
Accepted: October 11, 2012;
Published: February 09, 2013
How to cite this article
A. Norashikin, Z. Muta Harah and B. Japar Sidik, 2013. Intertidal Seaweeds and their Multi-life Forms. Journal of Fisheries and Aquatic Science, 8: 452-461.
DOI: 10.3923/jfas.2013.452.461
URL: https://scialert.net/abstract/?doi=jfas.2013.452.461
DOI: 10.3923/jfas.2013.452.461
URL: https://scialert.net/abstract/?doi=jfas.2013.452.461
INTRODUCTION
Seaweeds are the macroscopic marine algae that grow in the shallow coastal area. The majority of macroscopic marine algae or seaweeds are distinctly divided into three divisions; Chlorophyta, Phaeophyta and Rhodophyta (Dawson, 1966; Phang, 2006; Harah et al., 2007). Most seaweeds are growing in the shallow intertidal zone throughout the world (Rost et al., 1984; Mantri, 2005) but they can extend into the deeper subtidal zone as well. Seaweeds occurring in the intertidal zone are submersed and immersed periodically or aperiodically due to tides or irregularity occurring factors (Luning, 1990). The seaweed community occurrence in a particular intertidal shore are influenced by wide range of environmental factors such as nutrients, temperature and salinity (Chung et al., 2007), temperature, substratum and wave exposure (McQuaid and Branch, 1984), turbidity (Su et al., 2009) and salinity (Russell, 1988; Karsten, 2007).
Their presence in the intertidal areas are due to their ability to colonize solid substrates such as rock, dead corals and pebbles via holdfast structures for attachment (Akila and Jeyadoss, 2010). Okuda (2008) studied seaweed life forms and their interaction with others and how seaweeds molded in their habitats. Seaweed life forms can be classified into five types; epipelic; attached to mud and sand, epilithic; living on stones, rock, boulders or dead corals, epiphytic; attached to seagrass or other macroalgae, epizoic; attached to mollusc shells or polychaete and drift seaweed; loose lying macroalgae (Bold and Wayne, 1978; Den Hartog, 1979; Brouns and Heijs, 1991; Harah et al., 2006). The seaweed communities in the tropical areas, although present received less attention, due to them being non-visible as they are in the water that make them less explored and thus the information concerning the diversity between communities is still limited (Harah et al., 2006; Wong et al., 2012). In the present study, two intertidal sites with distinct landform and landscape characteristics have been selected to examine the diversity distribution and life forms of the seaweed communities. This study also examined seaweeds' structural adaptations that enable them to colonize the available hard and soft substrates in the two sites i.e., Teluk Kemang and Teluk Pelanduk, Port Dickson, Negeri Sembilan, Malaysia.
MATERIALS AND METHODS
Study site: Two distinct intertidal sites, Teluk Kemang (2°27'52.63"N, 101°50'52.12"E) and Teluk Pelanduk (2°25'7.85"N, 101°52'58.19"E) in Port Dickson, Negeri Sembilan, Malaysia were chosen for the study from November to December 2011 and March to May 2012 during low tide period. Teluk Kemang is a rocky outcrops area extending up to 50 m seaward during low tide, with gradual slope with live and dead corals interspersed with sand. Teluk Pelanduk, is a stretch of up to 150 m flat beach with mostly mud, sand and little rocks in between.
Field survey: A line transect method was used for collection of samples from intertidal and subtidal zones. At each site, line transects were laid perpendicular to the shoreline during the low tide period. The survey party move from the shore to the sea along the line transect of 100 m tape, recording all seaweed species, the nature of substratum, the point distant and representative plants were collected. These data were used for preparing the profile of each sampling site showing topography, landscape, seaweed species distribution and zonation.
Species identification: Seaweed samples collected were cleaned in situ with seawater to remove sand, debris and shells. Samples were placed in labelled plastic bags, preserved in 4% formalin-seawater solution and transferred to laboratory at Faculty of Agriculture, Universiti Putra Malaysia, Serdang, Selangor, Malaysia. Seaweed species identity was confirmed by referring to standard references such as Taylor (1960), Lewmanomont and Ogawa (1995), Ahmad (1995) and Trono Jr. (1997, 2004). Morphological characteristic of the seaweed collected was observed under dissecting and compound microscopes. Samples were finally preserved as herbarium specimens.
Water parameter analysis: Environmental factors such as water pH, salinity, Dissolve Oxygen (DO), conductivity, turbidity, temperature and salinity were measured in situ using DKK-TOA branded Multi Parameter Water Quality (MM: WQC-24). Water samples were fixed by adding HCl and placed in cooled ice chest for nutrient analyses for phosphate, nitrite, nitrate and ammonia in the laboratory using HACH spectrophotometer.
Data analysis: The similarity coefficient, seaweed’s generalist and specialist species between the two sites were obtained using Jaccard Similarity Community analysis (Jaccard, 1908) based on a presence-absence matrix following the formula; Cj = a/(a+b+c), where, Cj is the Jaccard similarity coefficient, a is number of species common to both sites, b is number of species in site B, but not in site A and c is number of species in site A, but not in site B.
RESULTS AND DISCUSSION
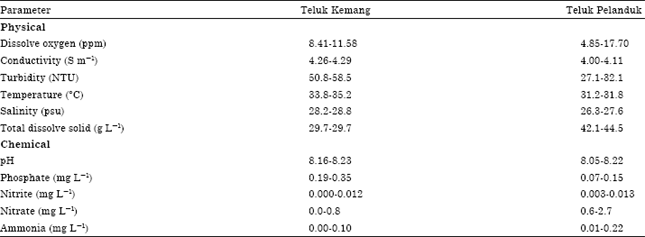
Seaweed community: Intertidal profiles of study sites are shown in Fig. 1 and 2. Seaweeds occurred in the stretch of 50-150 m from the shoreline toward the seas and particularly associated with hard substrates e.g., rocks. Seaweeds from the three divisions (Chlorophyta, Phaeophyta and Rhodophyta) were observed to be overlapping in distribution with no distinct zonation. According to Martin (2000) different environmental factors may vary with locality and govern the distribution of seaweed communities. Water quality parameters obtained in situ and based on laboratory chemical analyses is shown in Table 1. Nitrate content of water in Teluk Pelanduk ranges at 0.6-2.7 mg L-1 and Teluk Kemang ranges at 0.0-0.8 mg L-1 and in contrast turbidity of water at Teluk Kemang (50.8-58.5 NTU) and Teluk Pelanduk (27.1-32.1 NTU). Total Suspended Solid (TSS) affects the turbidity of the water. The higher the TSS, the more turbid is the water (Slingsby and Cook, 1986). Phang (1988) reported the sheltered west coast of Peninsular Malaysia is especially vulnerable to silts from land-based activities and the silts have been cited as one of the pollutants that affect the seaweeds’ survival. Large flexible or branched forms like Sargassum and Padina only occurred at Teluk Kemang and absent at Teluk Pelanduk (Table 2) are less susceptible as they are able to avoid smothering by silt. Seaweed species distribution at the sites reflected those that adapt or have adapted to the environmental conditions, particularly in utilizing the availability of the existing hard and soft substrates.
 | |
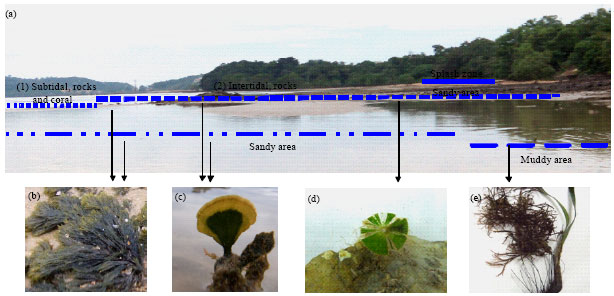
| Fig. 1(a-e): | (a) Coastal profile of Teluk Kemang showing the types of substrate and examples of seaweed species (b) Sargassum duplicatum, (c) Caulerpa microphysa, (d) Caulerpa racemosa and (e) Halimeda opuntia distributed in intertidal and subtidal areas |
 | |
| Fig. 2(a-e): | (a) Coastal profile of Teluk Pelanduk showing the types of substrate and examples of seaweed species (b) Cladophora prolifera, (c) Avrainvillea erecta, (d) Acetabularia major and (e) Gracilaria changii attach to Enhalus acoroides in the intertidal and subtidal areas |
| Table 1: | Environmental parameters of Teluk Kemang and Teluk Pelanduk |
 | |
A total of 43 species of seaweed were recorded, of which 36 species (16 Chlorophyta, 10 Phaeophyta, 10 Rhodophyta) at Teluk Kemang and 26 species (11 Chlorophyta, 6 Phaeophyta, 9 Rhodophyta) at Teluk Pelanduk (Table 2). At the two sites there were 9 generalist species e.g., Caulerpa prolifera, Ulva intestinalis and Dictyota dichotoma. Seventeen specialist species e.g., Caulerpa lentillifera, S. duplicatum and Laurencia papillosa at Teluk Kemang while 7 specialist species e.g., Acetabularia major, Avrainvillea erecta at Teluk Pelanduk (Table 2). Fewer species were recorded at Teluk Pelanduk compared to Teluk Kemang. This is attributed to its topography and landscape, although with longer intertidal area of unstable soft substrates e.g., mud and sand but the area lacks availability of stable hard substrates for seaweed colonization. The presence of different substrates rocks, sand, live and dead corals at Teluk Kemang widens the diversity of seaweeds that can adapt to these conditions (Trono Jr., 1986) and the stability of substratum obviously determines the species composition (Norton et al., 1981).
| Table 2: | Seaweed species and life forms in Teluk Kemang and Teluk Pelanduk |
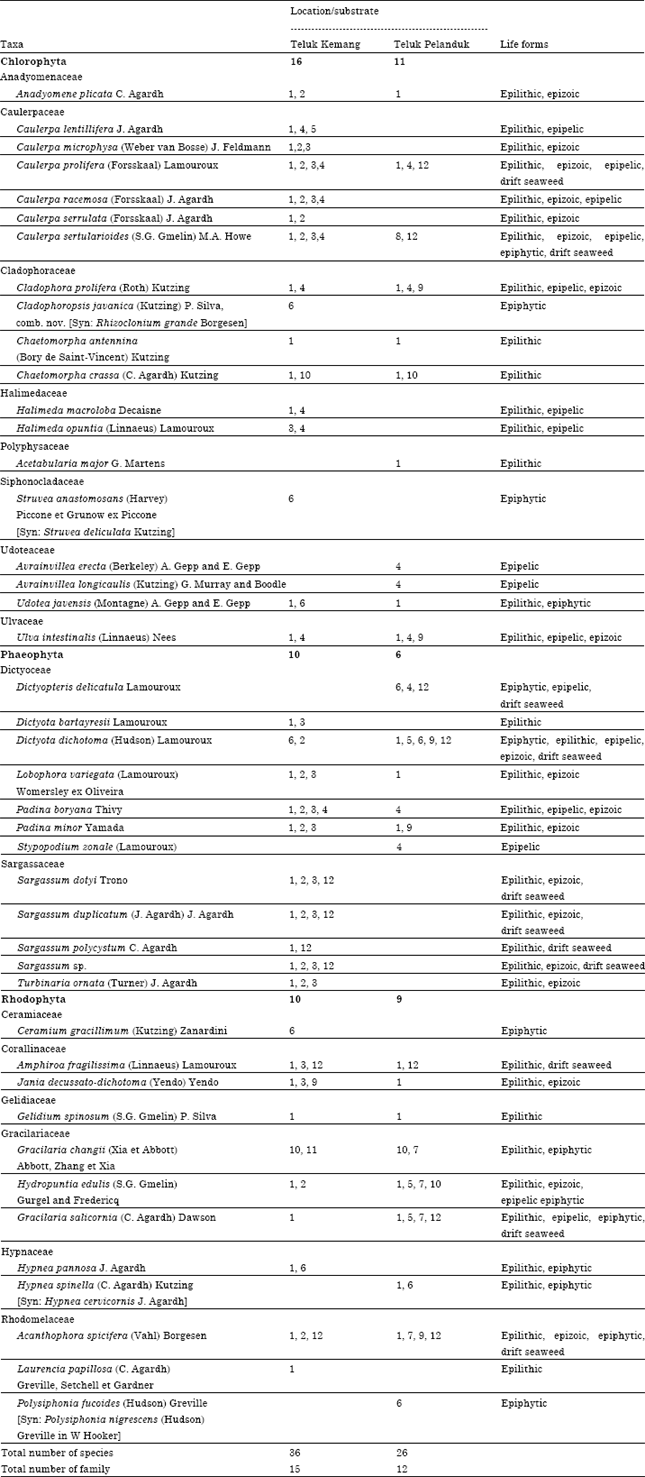 | |
| Substrate; 1: Rock, 2: Live coral, 3: Dead coral, 4: Sand, 5: Mud, 6: Other seaweed, 7: Seagrass, 8: Mangrove root, 9: Sea shell, 10: Rope, 11: Tyre, 12: Drift seaweed | |
Based on Jaccard community similarity analysis, a coefficient of 0.442 was obtained supporting the above differences that seaweed composition and diversity varied between the two areas.
Life forms of seaweed: Seaweeds were inhabiting diverse substrates from rocks, boulders, coral rubbles, dead corals to muddy and sandy areas. The seaweed life forms at Teluk Kemang were epilithic (45.71%), epizoic (24.29%), epipelic (12.86%), epiphytic (8.57%) and drift seaweed (8.57%).
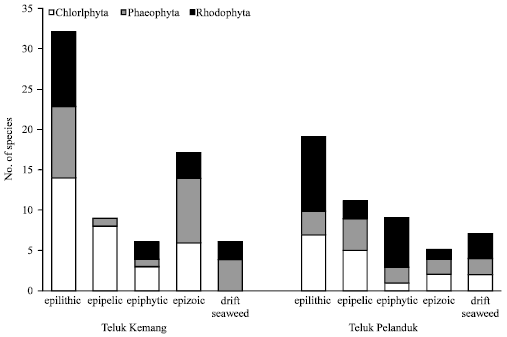 | |
| Fig. 3: | The multi-life forms based on seaweed divisions at Teluk Kemang and Teluk Pelanduk |
Those at Teluk Pelanduk were epilithic (38.46%), epipelic (21.15%), epiphytic (17.31%), drift seaweed (13.46%) and epizoic (9.62%). Epilithic seaweeds represent a big group for both areas with 32 species at Teluk Kemang and 20 species at Teluk Pelanduk (Fig. 3). Seaweed under this category utilised substrates ranging from rocks, live corals and dead corals to coral rubbles. Caulerpa lentillifera and C. sertularioides equipped with rhizoidal stolons were growing among live corals while Sargassum species, Turbinaria ornate possessed holdfasts that enabled them to establish on hard surfaces e.g., rocks, dead corals and coral rubbles. The epilithic seaweeds were also common in rocky areas e.g., Talibong island in Thailand (Prathep et al., 2008). Copejans et al. (1992) reported that most of the seaweeds at Gazi Bay, Kenya were attached to hard substrates such as rock and coral. Species such as Halimeda opuntia had rhizoids that allow them to attach tightly on the hard substrate. Live corals provide poor substrate for most seaweed colonisation and their sporelings. This is attributed to live corals capable of releasing antibiotic substances that inhibit the settlement of seaweed spores in the vicinity (Goh and Chou, 1992). Rhizophytic seaweed such as Avrainvillea species, Stypopodium zonale (at Teluk Pelanduk) and, Halimeda macroloba (at Teluk Kemang) colonized and grew in the bottom topography that constitutes soft substrates e.g., sand or mud is under the epipelic category. The manner of attachment to the substrates differs widely in seaweeds from a holdfast of single modified basal cell, to various kinds of penetrating or entangling rhizoid, multicellular, adherent discs, creeping stolons and massive clasping hapteres (Dawson, 1966). Several species occurred in two or more categories, since the same macroalgae utilised a range of substrates, e.g., Anadyomene plicata, Caulerpa microphysa, Cladophora prolifera can be epilithic and epizoic. Similarly Acanthophora spicifera, Amphiroa fragilissima and Gracilaria changii are epiphytic on other seaweeds or seagrass (e.g., Enhalus acoroides at Teluk Pelanduk). Simultaneously due to storms or heavy waves, they get detached as drift seaweeds which remain free or suspended among substrata.
Several seaweed species e.g., Gracilaria changii, Hydropuntia edulis and Chaetomorpha crassa utilized abandoned fishing lines and nets, ropes and car tyres for substrates.
The two sites under studied demonstrated diverse species composition and mode of attachment which reflected the multi-life forms that enabled them to colonise the marine environment. This is attributed to their adaptability to the availability of wide range of substrates ranging from sand to mud bottom, live corals, dead corals, coral rubbles (Norton et al., 1981; Fakoya et al., 2011) and marine plants. Some macroalgae are not attached to any substratum but remain free and still can survive in the environment.
CONCLUSION
This study revealed the seaweeds’ composition distribution, diversity and life form, of two contrasting intertidal areas at Teluk Kemang and Teluk Pelanduk. There was no discernible species horizontal zonation as seaweed species overlapped in their distribution. Seaweeds species were equipped with structure to colonize the available natural hard and soft substrates adaptations and their life forms were diverse. Abandoned drift objects (fishing nets and lines, ropes and tyres) introduced by human also formed the substrates for seaweed colonization.
ACKNOWLEDGMENT
We would to thank Vice-Chancellor, Universiti Putra Malaysia, for encouragement and facilities. This research is funded by Ministry of Science and Technology and Innovation Malaysia under Science Fund entitled “Utilization of aquatic macrophytes for papermaking”.
REFERENCES
- Chung, I.C., R.L. Hwang, S.H. Lin, T.M. Wu and J.Y. Wu et al., 2007. Nutrients, temperature and salinity as primary factors influencing the temporal dynamics of macroalgal abundance and assemblage structure on a reef of Du-Lang Bay in Taitung in southeastern Taiwan. Bot. Stud., 48: 419-433.
Direct Link - Fakoya, K.A., F.G. Owodeinde, S.L. Akintola, M.A. Adewolu, M.A. Abass and P.E. Ndimele, 2011. An exposition on potential seaweed resources for exploitation, culture and utilization in West Africa: A case study of Nigeria. J. Fish. Aquat. Sci., 6: 37-47.
CrossRef - Goh, N.K.C. and L.M. Chou, 1992. A Comparison of Benthic Life-form Characteristics of a Reef (Cyrene) Nearest to and a Reef (Raffles lighthouse) Furthest from Mainland Singapore. In: Marine Science: Living Coastal Resource, Chou, L.M. and C.R. Wilkinson (Eds.). Department of Zoology, National University, Singapore, pp: 55-61.
- Jaccard, P., 1908. Nouvelles rescherches sur la distribution florale. Bull. Soc. Sci. Naturale, 44: 223-270.
Direct Link - Karsten, U., 2007. Salinity tolerance of Arctic kelps from Spitsbergen. Phycol. Res., 55: 257-262.
CrossRefDirect Link - McQuaid, C.D. and G.M. Branch, 1984. Influence of sea temperature, substratum and wave exposure on rocky intertidal communities: An analysis of faunal and floral biomass. Mar. Ecol. Prog. Ser., 19: 145-151.
Direct Link - Harah, Z.M., B.J. Sidik, A. Raesah, S.A. Awing and O. Hisao, 2006. Marine macrophytes: Macroalgae species and life forms from Golden Beach, Similajau National Park, Bintulu, Sarawak, Malaysia. Coastal Mar. Sci., 30: 243-246.
Direct Link - Okuda, K., 2008. Coastal environment and seaweed-bed ecology in Japan. Kuroshioo Sci., 2: 15-20.
Direct Link - Phang, S.M., 2006. Seaweed resources in Malaysia: Current status and future prospects. Aquat. Ecosyst. Health Manag., 9: 185-202.
CrossRefDirect Link - Russell, G., 1988. The seaweed flora of a young semi-enclosed sea: The baltic. Salinity as a possible agent of flora divergence. Helgolande Mer. Res., 42: 243-250.
Direct Link - Su, W.C., I.C. Chung and T.M. Lee, 2009. Temporal dynamics of rocky-shore macroalgal assemblage structures in relation to coastal construction threats in Orchard Island (Taiwan): Impacts of turbidity and nutrients on the blooms of Galaxaura oblongata and a red alga-sponge symbiose Ceratodictyon/Haliclona. Kuroshio Sci., 3: 63-80.
- Wong, S.C., Z.M. Harah, B.J. Sidik and A.B. Arshad, 2012. Comparison of seaweed communities of the two rocky shores in Sarawak, Malaysia. Coastal Mar. Sci., 35: 78-84.
Direct Link