
Z. Muta Harah
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
B. Japar Sidik
Faculty of Agriculture and Food Sciences, Universiti Putra Malaysia, Bintulu Sarawak Campus, Nyabau Road, 97008 Bintulu, Sarawak, Malaysia
Journal of Fisheries and Aquatic Science
Year: 2013 | Volume: 8 | Issue: 3 | Page No.: 441-451







ABSTRACT
The Perhentian Island archipelago, Terengganu in east coast of Peninsular Malaysia is relatively in isolation from the mainland and poor accessibility has led to the paucity of observation on marine animals and plants e.g., seagrasses for the archipelago. Field surveys on seagrasses around Perhentian Besar Island and Perhentian Kecil Island was conducted for occurrence and distribution of seagrasses. Plants were sampled by direct collection during low tide or by snorkelling and SCUBA diving over four years (2007-2010) as part of the inventory of marine plants. Five species of seagrasses comprising the rare larger-bodied, Enhalus acoroides (L.f.) Royle and the small-bodied Halophila decipiens Ostenfeld, H. minor (Zoll.) den Hartog, H. ovalis (R. Brown) Hooker and Halodule pinifolia (Miki) den Hartog were recorded around the two islands. Enhalus acoroides grew in isolated patches at shallow depths of 0.5-1.5 m of the littoral zone usually exposed during the low spring tides at Tanjung Batu Lepir. Halophila decipiens, H. minor, H. ovalis and Halodule pinifolia grew as monospecific or mixed populations with no distinct zonation distributed at deeper depths of 4.6-12.0 m of the sublittoral zones at Channel in front of Department of Fisheries, Channel in front of Coco Hut and Kg. Tanjung Aur. Seagrass characterized by small-bodied with flaccid leaves e.g., Halophila decipiens, H. minor, H. ovalis and Halodule pinifolia are uniquely equipped for survival under extreme physical oceanographic due to seasonal north-east monsoons prevailing at Perhentian islands, making them less vulnerable to wave action and become detached. The morphological descriptions of the five species are also described.
PDF Abstract XML References Citation
Received: September 19, 2012;
Accepted: October 10, 2012;
Published: February 09, 2013
How to cite this article
Z. Muta Harah and B. Japar Sidik, 2013. Occurrence and Distribution of Seagrasses in Waters of Perhentian Island Archipelago, Malaysia. Journal of Fisheries and Aquatic Science, 8: 441-451.
DOI: 10.3923/jfas.2013.441.451
URL: https://scialert.net/abstract/?doi=jfas.2013.441.451
DOI: 10.3923/jfas.2013.441.451
URL: https://scialert.net/abstract/?doi=jfas.2013.441.451
INTRODUCTION
Seagrasses are unique amongst flowering plants as they are able to live submerge in the marine environment (Den Hartog, 1970). Apart from macroalgae (seaweeds) being the major autotrophic component of marine ecosystem (Penhale, 1977; Borum and Wium-Andersen, 1980), seagrasses are also important primary producer, providing refugia or nursery areas for many fishes and crustaceans (Bell and Pollard, 1989; Larkum et al., 1989; Poiner et al., 1987; Sasekumar et al., 1989) and as substratum for many epiphytes (Bujang et al., 2006). In our biogeographical studies, collections have been made of seagrass plants in coastal waters around the mainland Peninsular Malaysia. The distribution of seagrasses in Peninsular Malaysia has been detailed in various publications (Den Hartog, 1970; Japar Sidik, 1994; Bujang et al., 2006; Muta Harah et al., 2002a, 2003). The attention paid in the last decades has led to many records of species in areas where they had not been known before (Muta Harah et al., 1999, 2002a, b, 2003). In Malaysia, 15 species of seagrasses belonging to 8 genera, 3 families and 2 orders distributed over wide areas of Peninsular Malaysia (western, eastern and southern coasts) and East Malaysia have been recorded (Japar Sidik and Muta Harah, 2011).
In the east coast of Peninsular Malaysia in particular, the relative isolation and poor accessibility has led to the paucity of observation on marine plants for the region. Japar Sidik (1994) summarized the known distribution of seagrasses of Malaysia and noted this gap. The existing studies around off-shore islands e.g., Redang Island (Muta Harah et al., 2003) and Tinggi Island (Bujang et al., 2006) gave information on the occurrence and distribution for small-leaved species such as H. decipiens, Halodule pinifolia, as well as the bigger-leaved species, Cymodocea serrulata, C. rotundata and Syringodium isoetifolium. However, areas such as Perhentian Island archipelago of the east coast region remains unsurveyed for marine macrophytes e.g., seagrasses. This study collates species records and sporadic observation, providing occurrence, distribution and prevailing habitat information for the marine macrophytes found and their morphological characteristics described.
MATERIALS AND METHODS
Study area description: The Perhentian Island archipelago comprised of islands of Perhentian Besar, Perhentian Kecil and Susu Dara off the coast of Terengganu (Fig. 1).
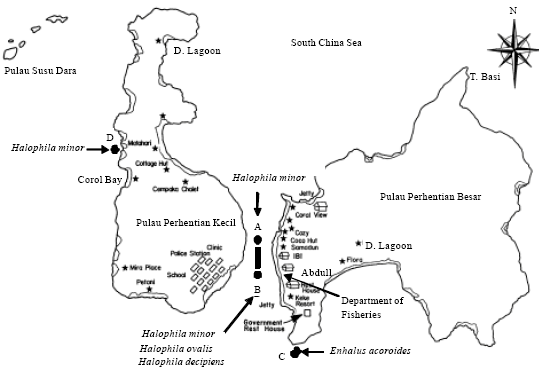 | |
| Fig. 1: | The location of seagrass species in waters of Perhentian Island Archipelago, A: Channel in front of Coco Hut, B: Channel in front of Department of Fisheries, C: Tanjung Batu Lepir, D: Kg. Tanjung Aur (Rajawali) |
The coastlines consist of sandy beaches interrupted by stretches of rock outcrops. Perhentian islands have more prominent rock formation but generally low lying and surrounded by a shallow coastal waters. Adjacent to the sandy beaches and sometimes the rocky outcrops in the littoral and sublittoral zones are seagrass patches and meadows. Their presence and distribution are dependent on the available and local prevailing environmental conditions.
Field surveys: Field surveys were undertaken in April 2007 to June 2010 along the coastal areas of Perhentian Besar and Perhentian Kecil islands by SCUBA diving. Four locations (Fig. 1) of seagrass were identified; Tanjung Batu Lepir, Channel in front of Department of Fishery, Channel in front of Coral View and Kg. Tanjung Aur near to Cord Bay (Rajawali). Botanical surveys at each location were carried out with respect to occurrence, approximate cover, abundance (by visual estimation) and associated biota (macroalgae). For a particular location, habitat type, depth was recorded and then seagrass species was listed and collected at random. They were preserved in buffered 4% formalin in saline water (Menez et al., 1983) for laboratory investigation.
Seagrass identification: The identity of the seagrass was ascertained by referring to Den Hartog, (1970), Sidik et al. (1995), Japar Sidik et al. (1999) and Kuo (2000). Vegetative (leaf length, leaf width, space between two cross veins, space between intramarginal vein and blade margin, petiole length, rhizome diameter and rhizome internode length) and reproductive (male and female flowers and fruit if available) dimensions were recorded.
RESULTS
Occurrence and distribution: The waters of Perhentian Island archipelago with little variation in salinity of 29-32 psu (practical salinity unit) and temperature 28-31°C harboured five species of seagrasses; Enhalus acoroides (L.f.) Royle, Halophila decipiens Ostenfeld, H. minor (Zoll.) den Hartog, H. ovalis (R. Brown) Hooker and Halodule pinifolia (Miki) den Hartog. With the exception of Enhalus acoroides, all species were small-bodied seagrasses.
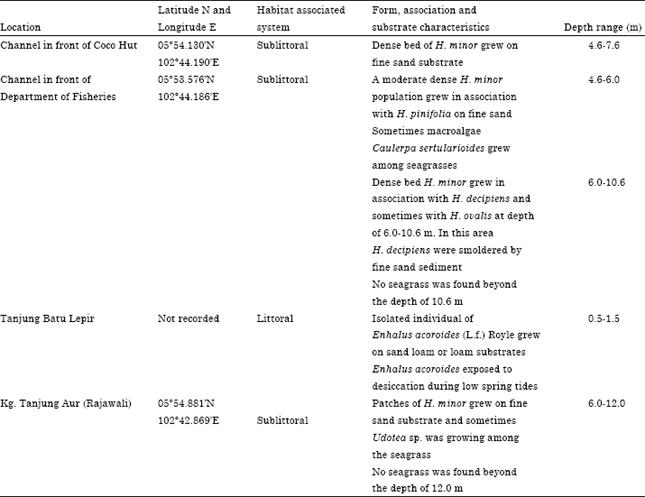
Except for the rare E. acoroides growing in the littoral zone on sand to sand loam substrates at Tanjung Batu Lepir, the other four species were thriving in the sublittoral zone with sand substratum (Table 1, Fig. 2). Isolated individuals of E. acoroides inhabited the shallow depths of 0.5-1.5 m usually exposed during the low spring tides. Patches to dense populations of Halophila decipiens, H. minor, H. ovalis and Halodule pinifolia inhabited the sea bottom at variable deeper depths of 4.6-12.0 m.
Dense bed of monospecific H. minor grew on fine sand substrate at Channel in front of Coco Hut and sometimes growing with seaweed, Udotea sp. at Kg. Tanjung Aur (Rajawali). However, at Channel in front of the Department of Fisheries it grew as a moderate dense bed in association with H. pinifolia and H. decipiens and sometimes with H. ovalis (Table 1). Halophila ovalis, H. decipiens and Halodule pinifolia were found only at Channel in front of Department of Fisheries.
Description of seagrasses: Based on the dimension analysis of vegetative structures e.g., leaf length, leaf width, space between two cross veins, space between intramarginal vein and blade margin, rhizome internode length, rhizome diameter, the descriptions of the Perhentian Island archipelago seagrasses are given below.
| Table 1: | Demographic characteristics of seagrasses in waters of Perhentian Island Archipelago |
 | |
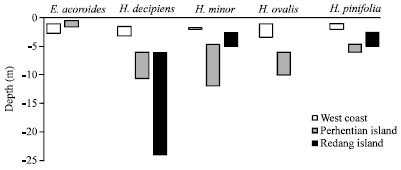 | |
| Fig. 2: | Comparison of depth distribution of 5 seagrass species in the west coast and east (Perhentian and Redang islands) |
Enhalus acoroides (Fig. 3a), the local rare species grew as isolated individual plants in sand-loam substrates of littoral area of Tanjung Batu Lepir. Enhalus acoroides is a large-bodied monomorphic species consisting of tough rhizome creeping coarsely under sediment and the shoot with leaves. Attached to the rhizome are 2-5 strap-like leaf blades with many parallel nerves and round leaf tip. The young leaf is light green in color, protected between two or more older leaf blades.
 | |
| Fig. 3(a-g): | Seagrass species from Perhentian Island Archipelago. (a) Enhalus acoroides from Tanjung Batu Lepir, (b) Halophila decipiens from Channel in front of Department of Fisheries, (c) Halophila decipiens male flower (mf) and female flower (ff) on the same shoot, (d) Halophila decipiens fruit, (e) Halophila minor from Channel in front of Coco Hut, Channel in front of Department of Fisheries and Kg. Tanjung Aur (Rajawali), (f) Halophila ovalis, a small population growing with H. minor and H. decipiens at Channel in front of Department of Fisheries and (g) Halodule pinifolia grew in association with H. minor at Channel in front of Department of Fisheries |
This is to protect the young leaf from getting damage by wave or grazed by other organisms. The mature leaves are dark green in color and long, ranged from 71.00-103.50 cm in length; 0.90-2.00 cm in width. The whole part of the rhizome is covered with persistent black bristles for protection. These black bristles are from old degraded leaf sheath forming the brown or black colored bristles. The diameter of rhizomes varies from 1.27-2.29 cm. Root of the E. acoroides is pale yellow in color and arise from the lower surface of the rhizome. It is spongy, soft, cord in shape, unbranch and numerous without root hairs. The length and diameter of root varies from 7.70-27.10 cm and 0.22-0.56 cm, respectively. No flowers were observed. This species tend to be relatively high desiccation tolerance due largely to its leaves which lie overlapped on the sediment surface when the tide is low.
Halophila decipiens is a monoecious plant (Fig. 3b) with a creeping fragile narrow rhizome, 0.40-1.00 mm thick, rhizome internode length, 4.00-25.50 mm. One unbranched root with numerous root hairs is present below each erect shoot. Erect shoot is up to 2.50 mm long arising from each node, bearing a pair of leaves. The erect stem and the immediate rhizome are enveloped by a pair of obovate, keeled, transparent scales, with short stiff hairs on the margin and the outside, upper scale, 2.00-3.50x1.80-2.68 mm, lower scale, 2.00-3.80x1.30-2.30 mm. Leaf blade is pale green in colour, distinctly elliptic, obtuse at the apex, margin finely serrated, 10.50-17.00 mm long, 4.00-7.00 mm wide. The mid-vein is conspicuous, united apically with intramarginal veins, cross veins 5-10 pairs, sub-opposite to alternate, occasionally forked. The adaxial and abaxial leaf surfaces possessed dense, short, stiff unicellular hairs, almost evenly distributed; petioles slender, 3.00-8.00 mm long. Inflorescence, a pair of flowers on the same or lateral erect shoots; one male and one female flower, subtended by 2 overlapping scarious, transparent spathes with finely serrulate margins, 1.80-3.80 mm long. A male flower comprised of a whitish pedicel, 1.30-3.65 mm long; three whitish, oblong-elliptic, convex tepals, 0.70-1.00x0.32-0.50 mm and three fused anthers, 0.35-0.40 mm long (Fig. 3c). A female flower comprised of a sessile, ovoid ovary, 0.50-1.10x0.45-0.70 mm; a hypanthium 0.40-1.80 mm long; three styles, with glandular inner surfaces, of unequal length of 0.90-3.80 mm long (Fig. 3c). Fruits broadly ellipsoid, 1.00-2.10 mm long, 0.50-1.50 mm wide (Fig. 3d). Seeds 20-25, ellipsoid to spherical, bluntly beaked at both ends, testa reticulate.
Halophila minor is a petiolated seagrass (Fig. 3e) with creeping horizontal rhizome, 0.80-1.60 mm in diameter, rhizome internode length, 10.80-50.0 mm long, an unbranched root with numerous root hairs at each node. Scales two; transparent, convex or folded, suborbicular to transversely elliptic, acute, rounded or emarginate at the apex, auriculate at the base. The scale is 2.60-5.20x2.80-5.80 mm. Leaves in pairs, with petiole, 4.00-35.10 mm long. Leaf blade ovate or obovate, green, entire; apex rounded; base obtuse or shortly cuneate and contracted near the petiole, 11.00-19.70 mm in length and 5.90-12.10 mm in width, with a L:W ratio of 1.37-2.08:1; cross veins 7-11 pairs, space between two cross veins, 0.46-2.55 mm, space between intramarginal vein and blade margin narrow, 0.18-0.36 mm. Male and female flowers were not found.
Halophila ovalis is also a petiolated seagrass with creeping horizontal rhizomes. Rhizomes up to 1.57 mm in diameter with a single root at each node (Fig. 3f). Two petioles attached to the node are almost equal in length, ranged 14.20-35.90 mm. Leaf blade 18.20-31.45 mm in length and 11.30-17.05 mm in width. Cross vein 15 to 22 pairs per leaf, often forked, joining the intramarginal nerves. Nodal region also bears a pair of scale leaf, transparent, suborbicular or obovate, convex or folded with the length 2.80-5.70 mm and width 2.80-6.50 mm. One is sheathing the rhizome and another sheathing the leaf petioles. From our collection, only male flowers were observed. A male flower consists of three tepals; 3.57-4.14 mm long, up to 1.90 mm in width, white with a dark strip in the middle, transparent, convex and elliptic; pedicel, thin and whitish, 12.57 mm long enveloped by a pair of spathes, 2 mm wide and 3.43 mm long. Three oblong anthers were present, 2.86-3.28 mm long.
Halodule pinifolia had a creeping rhizome, 0.48-1.06 mm in thickness with 1-3 roots per node (Fig. 3g). The rhizome internode measured 1.97- 28.00 mm in length. Erect stem, short varies from 1.52-20.90 mm long bearing 1-4 delicate linear leaf blade, dark green in colour. Length of the leaves ranged from 39.00-97.50 mm and its width ranged 0.66-1.00 mm. The leaf tips are obtuse with more or less irregular serratures; lateral teeth are temporary developed or totally absent. Midrib conspicuous, both ends in a very small tooth. The leaf sheath is short, with the range of 1.72-19.40 mm. Male and female flowers were absent.
DISCUSSION
Occurrence and distribution: In this present study five species of seagrasses such as E. acoroides, Halophila decipiens, H. minor, H. ovalis and Halodule pinifolia were recorded around the two islands; Perhentian Besar Island and Perhentian Kecil Island, Terengganu. In contrast, 12 species are known for the west coast, including several larger-bodied, wide-leaved seagrasses such as Enhalus acoroides, Thalassia hemprichii (Ehrenb.) Aschers., Cymodocea serrulata (R. Br.) Aschers. and Magnus and C. rotundata Ehrenb. and Hempr. ex Aschers (Bujang et al., 2006). The relatively low species of larger-bodied seagrass is an indication of stressful conditions prevailing at Perhentian islands. Extreme physical oceanographic conditions prevalent in the east coast of Peninsular Malaysia, due to seasonal north-east monsoons occurring from November to January (Zelina et al., 2000), placed a high demand on the seagrass community e.g., at Perhentian and Redang Island archipelagos (Muta Harah et al., 2003). The physical features in particular e.g., waves fluctuate along with the water quality (e.g., sediments) conditions during monsoons, create a dynamic and fluctuating environment. Only seagrass characterized by small-bodied with flaccid leaves e.g., Halophila decipiens, H. minor, H. ovalis and Halodule pinifolia are uniquely equipped for survival under such extreme stressed conditions, making them less vulnerable to wave action and become detached. The seasonal physical oceanographic conditions may also prevent the establishment of larger-bodied seagrass common in the relatively calm, sheltered areas of west coast of Peninsular Malaysia.
Halophila decipiens, H. minor, H. ovalis and Halodule pinifolia are prolific seed producers e.g., H. decipiens rely completely on seed production (Kuo et al., 1995; Kirkman and Kuo, 1996) to maintain population in seasonally fluctuating environment (Kenworthy, 1992). Halophila ovalis and H. decipiens were in flowering and fruiting during the several samplings. Flowering and seed production would be an ideal mechanism to survive in deeper water under monsoonal conditions. Reproduction by seeds may also enhance survival in deeper water by allowing seagrass to persist through period of poor monsoonal water quality and unfavourable environmental conditions as temporary seed bank in the sediment (Kenworthy et al., 1989).
The ability of H. ovalis growing in sublittoral area showed that it has a lower minimum light requirement than other seagrasses (Kirkman and Kuo, 1996). In addition, H. ovalis was able to maintain high growth rates at low light levels (Hillman et al., 1995). Another conspicuous seagrass was H. decipiens. Sidik et al. (1995) documented its occurrence at shallow depth of -1.5 to -3.0 m MSL in turbid marine waters of Teluk Kemang, Negri Sembilan in the west coast of Peninsular Malaysia. Halophila decipiens has been recognised as a deep-water species usually occurring in waters between 10 m and 30 m depths (Den Hartog, 1970; Muta Harah et al., 2003). The seagrass, H. pinifolia was also observed in the sublittoral area. Phillips and Menez (1988) accredited this species to being tolerant to a wide range of depths. In a depth limit report compiled on 31 seagrass species, H. pinifolia can extend into waters of at least 14 m depth (Duarte, 1991). Coppejans et al. (1992) also reported that H. pinifolia could be found not only in littoral waters but also at considerable depths.
The present study revealed that seagrass species grew as monospecific or mixed populations with no distinct zonation or depth distribution. Halophila minor was more predominant (in terms of overall bottom coverage, frequency of occurrence) occurring at three locations; Channel in front of Coco Hut, Channel in front of the Department of Fisheries and Kg. Tanjung Aur. It grew in mixed meadows of up to three seagrasses (e.g., H. ovalis, H. decipiens, H. pinifolia) coexisted. Halophila ovalis, H. decipiens and Halodule pinifolia were found only at Channel in front of Department of Fisheries. No other seagrass species such as Halodule uninervis which is common in the east coast e.g., at Tengah, Besar, Tinggi islands down to Sibu island (Japar Sidik, 1994) was encountered. The range of depth limit of 4.6-6.0 m for H. pinifolia, H. minor and H. ovalis were in contrast to their shallow distribution in west coast of Peninsular Malaysia (Japar Sidik et al., 1999; Japar et al., 2001). In clear water, H. decipiens thrive at depth of 8-18 m at Sulug Island, Sabah, Malaysia (Japar Sidik et al., 1997) and 6-24 m at Redang Island (Muta Harah et al., 2003). Elsewhere around the world it occurred at similar depth or deeper and on a variety of substrates; at depth of 19-23 m on clayed sand at Tenerife, Canary Islands (Gil-Rodriquez et al., 1982), 11-23 m depth on soft mud with fine sand and silty sediments in the southern Philippines (Menez et al., 1983), 15-18 m depth on flat sandy coralline at Okinawa Island (Kuo et al., 1995), 58 m depth on carbonate mud in Northeastern, Australia (Long et al., 1989). In turbid water it was found at 2 m depth on muddy coral sand at Tuticorin, Southern India (Parthasarathy et al., 1988) and 1.5 m at Teluk Kemang, Port Dickson on sand-mud substrates Sidik et al. (1995). Altered depth of H. decipiens may be the ‘indicator’ on water quality changes that caused changes in light penetration and consequently affected its depth distribution pattern.
Description of seagrasses: The H. minor identified in this study is based on the recent taxonomic reexamination of H. ovata-H. minor complex, where H. ovata and H. minor are under separate distinct species Kuo (2000). The previous classification by Den Hartog (1970) and Sachet and Fosberg (1973) assumed that H. ovata and H. minor are identical. Kuo (2000) in his examination of other materials reported that H. minor is widely but disjunctively distributed in the tropical Indian Ocean and Western Pacific Ocean, including Kenya, India, Malaysia and Northern Australia while H. ovata has been collected from Saipan, Guam in the northern Mariana Islands, Yap or Micronesica Islands and Manila Bay in the Philippines in the western Pacific Ocean.
In Malaysia, H. pinifolia have been classified as long leaved variant (mean leaf length range 83.91-102.52 mm, leaf length range 60-166 mm) and short leaved variant (mean leaf length range 49.70-57.43 mm, leaf length range 40-80 mm). The Perhentian islands population belongs to the H. pinifolia short leaved variant (Japar Sidik et al., 1999). This species can be mistaken for H. uninervis narrow leaved variant where mature leaf-tips are tridentate with the median tooth as long as or even shorter or longer than the lateral. It is easily distinguished from H. uninervis broad leaved species from other areas in Malaysia, where the leaf blades are generally wider (mean leaf width ranged 2.16-3.20 mm, leaf width range 1.5-4.0 mm) and leaf-tips with an obtusely rounded or pointed median tooth in which the midrib ends, usually as long as and rarely longer than the 2 lateral teeth Japar Sidik et al. (1999).
Based on our studies on seagrass diversity and distribution (Bujang et al., 2006), a very broad distinction can be made between the seagrass distribution of the east and west coasts. The types of available habitat and prevailing environmental characteristics along the east and west coasts probably explain these differences. On the west coast larger-bodied, wide-leaved seagrass e.g., T. hemprichii, C. serrulata and C. rotundata are common while the east coast with the exception of E. acoroides, are limited to the small-bodied seagrass e.g., H. decipiens, H. minor, H. ovalis and H. pinifolia. The west coast of Peninsular Malaysia does not generally experience strong wave action, whereas the east coast is exposed annually to the seasonally north-east monsoon from November to January (Zelina et al., 2000). Clarity of water and sufficient light irradiance during the off monsoon season play a significant role in the depth distribution of the seagrasses. Coastal waters in the west coast of Peninsular Malaysia are often turbid or high in suspended solids that limit the depth at which most seagrass grow. This is reflected in seagrass communities along the west coast which are generally found inhabiting the shallow waters at depth less than 4.0 m (Fig. 2). In terms of species diversity distribution with depth e.g., seagrass area stretching from Mandapam to Thondi regions of Palk Bay, Southeastern India, it has been reported that species distribution and composition is reduced from nearshore (0-2.0 m depth) to middle zone (2.1-4.0 m depth) and offshore (4.1-7.0 m depth) (Manikandan et al., 2011). They attributed the differences among others are due to favourable salinity, less depth and less sedimentation in nearshore. Seagrasses in the east coast particularly those around Perhentian and Redang Island archipelagos however extend to deeper areas, greater than 4 m to 24.0 m where the water is clear. Seagrasses will colonize greater depth if the water is clear (Duarte, 1991).
CONCLUSION
The five species of seagrasses recorded from Perhentian Island archipelago, east coast Peninsular Malaysia comprised the rare larger-bodied Enhalus acoroides and, small-bodied Halophila decipiens, H. minor, H. ovalis and Halodule pinifolia. The small-bodied species thrived in sublittoral and one species (Enhalus acoroides) at littoral zones. The seagrasses at sublittoral grew at deeper depth of 4.6-12 m on sand to sand loam substrates and they formed either monospecific or mixed populations. Compared to the west coast Peninsular Malaysia, the common seagrasses are the larger-bodied e.g., T. hemprichii, C. serrulata and C. rotundata and are generally of mixed population inhabiting the sandy to muddy substrates at shallow water depth of less than 4.0 m.
ACKNOWLEDGMENT
We would like to thank the Vice-Chancellor, Universiti Putra Malaysia, for encouragement and facilities. The research fund provided by the Ministry of Science, Technology and Environment Malaysia, under the ‘Intensification of Research in Priority Areas’ programme is acknowledged.
REFERENCES
- Bell, J.D. and D.A. Pollard, 1989. Ecology of Fish Assemblages and Fisheries Associated with Seagrasses. In: A Treatise on Seagrasses with Special Reference to the Australian Region, Larkum, A.W.D., A.J. McComb, S.A. Shepherd (Eds.). Elsevier Scientific Publishers, Amsterdam, The Netherlands, pp: 565-609.
- Coppejans, E., H. Beeckman and M. de Wit, 1992. The seagrass and associated macroalgal vegetation of Gazi Bay (Kenya). Hydrobiologia, 247: 59-75.
CrossRef - Gil-Rodriquez, M.C., J. Afonso-Carillo and W.W. de la Torre, 1982. Occurrence of Halophila decipiens ostenfeld on Tenerife, Canary Islands. Aquat. Bot., 12: 205-207.
CrossRef - Bujang, J.S., M.H. Zakaria and A. Arshad, 2006. Distribution and significance of seagrass ecosystems in Malaysia. Aquat. Ecosyst. Health Manage., 9: 203-214.
CrossRefDirect Link - Japar, S.B., Z.M. Harah, Z. Kanamoto and A.M. Pauzi, 2001. Seagrass Communities of the Straits of Malacca. In: Aquatic Resource and Environmental Studies of the Straits of Malacca: Current Research and Reviews, Japar, S.B., A. Arshad, S.G. Tan, S.K. Daud, H.A. Jambari and S. Sugiyama (Eds.). Malacca Straits Research and Development Centre, Serdang, Malaysia, pp: 81-98.
- Japar Sidik, B., Z. Muta Harah, A.M. Pauzi and M. Suleika, 1999. Halodule species from Malaysia-distribution and morphological variation. Aquat. Bot., 65: 33-45.
CrossRef - Kenworthy, W.J., C.A. Currin, M.S. Fonseca and G. Smith, 1989. Production, decomposition and heterotrophic utilization of the seagrass Halophila decipiens in a submarine canyon. Mar. Ecol. Prog. Ser., 51: 277-290.
Direct Link - Kuo, J., Z. Kanamoto, T. Toma and M. Nishihira, 1995. Occurrence of Halophila decipiens Ostenfeld (Hydrocharitaceae) in Okinawa Island, Japan. Aquat. Bot., 51: 329-334.
CrossRef - Manikandan, S., S. Ganesapandian and K. Parthiban, 2011. Distribution and zonation of seagrasses in the palk bay, Southeastern India. J. Fish. Aquat. Sci., 6: 178-185.
CrossRefDirect Link - Muta Harah, Z., B. Japar Sidik and O. Hishamuddin, 1999. Flowering, fruiting and seedling of Halophila beccarii Aschers. (Hydrocharitaceae) from Malaysia. Aquat. Bot., 65: 199-207.
CrossRef - Muta Harah, Z., B. Japar Sidik and A. Arshad, 2002. Flowering, fruiting and seedling of annual Halophila beccarii Aschers in peninsular Malaysia. Bull. Mar. Sci., 71: 1199-1205.
Direct Link - Muta Harah, Z., B. Japar Sidik and A.R. Fazrullah Rizally, 2003. Occurrence and morphological description of seagrasses from Pulau Redang, Terengganu, Malaysia. J. Teknol., 38: 29-39.
Direct Link - Sidik, B.J., A. Arshad, O. Hishamuddin and S. Bahar, 1995. Halophila decipiens Ostenfeld: A new record of seagrass for Malaysia. Aquat. Bot., 52: 151-154.
CrossRef - Poiner, I.R., D.J. Staples and R. Kenyon, 1987. Seagrass communities of the Gulf of Carpentaria, Australia. Aust. J. Mar. Freshwater Res., 38: 121-131.
CrossRef - Sasekumar, A., M.U. Charles Leh, V.C. Chong, D. Rebecca and M.L. Audery, 1989. The Sungai Pulai (Johor): A unique mangrove estuary. Proceedings of the 12th Annual Seminar of the Malaysian Society of Marine Sciences, November 18, 1989, Institute for Advanced Studies, University of Malaya, Kuala Lumpur, Malaysia, pp: 191-211.