
S. Shahbudin
Institute of Oceanography and Maritime Studies, Kulliyyah of Science, International Islamic University Malaysia, Jalan Istana, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
K.C.A. Jalal
Institute of Oceanography and Maritime Studies, Kulliyyah of Science, International Islamic University Malaysia, Jalan Istana, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
Y. Kamaruzzaman
Institute of Oceanography and Maritime Studies, Kulliyyah of Science, International Islamic University Malaysia, Jalan Istana, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
N. Mohammad- Noor
Borneo Marine Research Institute, Universiti Malaysia Sabah,Locked Bag 2073, 88999 Kota Kinabalu, Sabah, Malaysia
T. Chit Dah
Borneo Marine Research Institute, Universiti Malaysia Sabah,Locked Bag 2073, 88999 Kota Kinabalu, Sabah, Malaysia
B. Akbar John
Institute of Oceanography and Maritime Studies, Kulliyyah of Science, International Islamic University Malaysia, Jalan Istana, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
Journal of Fisheries and Aquatic Science
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 85-92







ABSTRACT
Artificial seagrass plays an important role for the distribution of benthic organisms and fishes. Based on this perspective a study was conducted in the Sepanggar Bay located in the Northern part of Kota Kinabalu, Malaysia from November 2006 to February 2007 to determine the potentiality of artificial seagrass as an alternative habitat for marine fishes. The artificial seagrass area is located approximately 100 m from the coastline at the depth between 6 and 10 m and made up of Dorken Advance Engineer Rubber which was tightened on a square 1 x 3 m metal mat. The sampling process was done by capturing photos and video recording. The picture of the fishes was taken as many as possible to increase the accuracy and precision of fish identification. A total of 497 fish species belong to 17 families were observed in 5 sampling periods throughout the study. Out of 17 families, Pomacentridae was dominant with 25.55% (127 individuals) followed by caesionidae with 22.13% (110 individuals). It was observed that Caesio sp. from a family Caesionidae was dominant genus in the artificial seagrass area, followed by Chromis sp. and Pomacentrus sp. from a family Pomacentridae. Overall Shannon diversity (H’) value of 2.5315 indicates artificial sea grass could play a vital role as a nutrient rich habitat for marine fishes.
PDF Abstract XML References Citation
Received: April 01, 2010;
Accepted: May 29, 2010;
Published: July 27, 2010
How to cite this article
S. Shahbudin, K.C.A. Jalal, Y. Kamaruzzaman, N. Mohammad- Noor, T. Chit Dah and B. Akbar John, 2011. Artificial Seagrass: A Habitat for Marine Fishes. Journal of Fisheries and Aquatic Science, 6: 85-92.
DOI: 10.3923/jfas.2011.85.92
URL: https://scialert.net/abstract/?doi=jfas.2011.85.92
DOI: 10.3923/jfas.2011.85.92
URL: https://scialert.net/abstract/?doi=jfas.2011.85.92
INTRODUCTION
The artificial seagrass is familiar as an alternative method to replace the destroyed natural seagrass ecosystem in many countries. It has been widely used as a soft engineering method to protect shoreline from erosion and as an alternative marine habitat for various marine organisms. Besides the ability to prevent coastal erosion, it also provide nursery ground for juveniles, habitat for small fishes, food for grazers and hiding place for small fishes thus playing major role in maintaining balance in marine environment. They stabilize bottom sediments with their dense roots and rhizomes that form a secure mat. This sediment stabilization and erosion prevention is especially important during storms and hurricanes that often threaten the coastal areas (Fairhurst and Graham, 2003).
Seagrass beds offer important feeding sites for fish (Weinstein and Heck, 1977; Heck and Orth, 1980) due to the high invertebrate biomass they support (De Troch et al., 2003; Nakamura and Sano, 2005). Seagrass fish assemblages are commonly dominated by fish that feed on abundant small crustaceans (Nakamura et al., 2003). Food availability has been found to explain the distribution of fish within seagrass beds (Grenouillet et al., 2002). Khalaf and Kochzius (2002) found that 79.9% of fish in seagrass area were planktivores, 5.5% fed on both fish and invertebrates, 5% were omnivorous and only 2.5% fed exclusively on invertebrates. Although several studies (e.g., Marasabessy and Hukom, 1989; Dolar, 1991) have reported on the diversity and abundance of seagrass fish assemblages within SE Asia their trophic structure remains undocumented. Furthermore it is also efficient in removing dissolved nutrients from waters that often enter coastal waters as a result of runoff from the land (Duffy, 2006). They trap fine sediments and particles from both land and water. The removal of sediments and nutrients from the water results in high water clarity and nutrient-poor waters required for the survival of coral reefs. Despite of playing important roles in marine environment, seagrass ecosystem has been degraded in many ways especially by human activities (Lee et al., 2001; De Silva et al., 1999). Nevertheless, some of the functions of natural seagrass cannot be established by artificial seagrass. Artificial seagrass cannot produce gases to increase the gas regulation in the water. Artificial seagrass does not have real roots as natural seagrass to absorb nutrients and consolidate the sand structure. The successiveness of seagrass to attract fish inside of it can play the vital role to increase the fish population in the area (Adams et al., 2004). Among the fishes, the commercial fish also inhabits in the seagrass meadow area, indirectly boosting the local fishery economics.
Realizing the study on artificial seagrass in Malaysia is still scanty, this study was aimed to determine the effectiveness of artificial seagrass as a marine habitat towards the conservation of marine fishes.
MATERIALS AND METHODS
Sampling sites: This study has been conducted in Sepanggar Bay, Northern part of Kota Kinabalu (Fig. 1). The data was collected between November, 2006 and February, 2007. The artificial seagrass area located approximately 100 m offshore from the coastline. The depth at the artificial seagrass area varied between 6 and 10 m. The artificial seagrass units are made by Dorken Advance Engineer Rubber. They were tightened on a square 1 x 3 m metal mat. The metal mat was submerged with six heavy breaks surrounding. The length of each artificial seagrass unit was 1 m where the water absorption was less than 0.3% and it was non-toxic material.
During sampling, the weather and time were the vital factors to be concerned. The sampling was carried out during a sunny day between 9 o’clock in the morning until 3.00 o’clock in the afternoon. This period receives optimum sunlight and provides the best visibility as stated by other marine biologists (Sogard and Able, 1994; Bell et al., 1985). The sampling process was done by capturing photo and video recording (Beck et al., 2001; Heck et al., 2003; Hemminga and Duarte, 2000). The picture of the fishes was taken as many as possible by Self Contained Underwater Breathing Apparatus (SCUBA) divers to increase the accuracy and precision of fish identification. Video recording was used to record the fish activities in the artificial seagrass area. The morphological characteristics (e.g., colour, body shape, fins shape, snout type) and the behavior of the fishes in the artificial seagrass area were recorded on underwater water-proof paper as followed the method (Barber et al., 1979) and the diversity of fishes was analyzed using Shanon diversity (Shannon and Weaver, 1949).
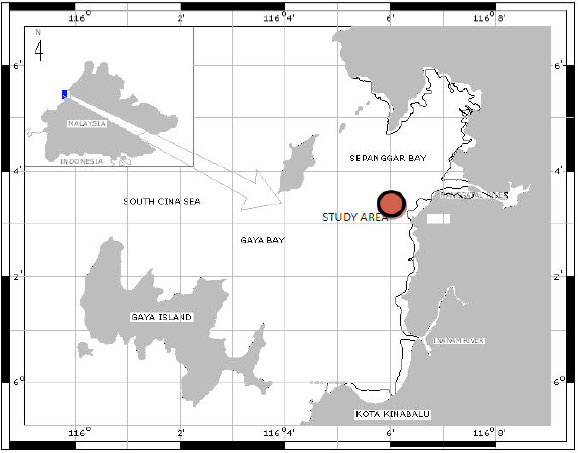 | |
| Fig. 1: | Location of the study area |
RESULTS
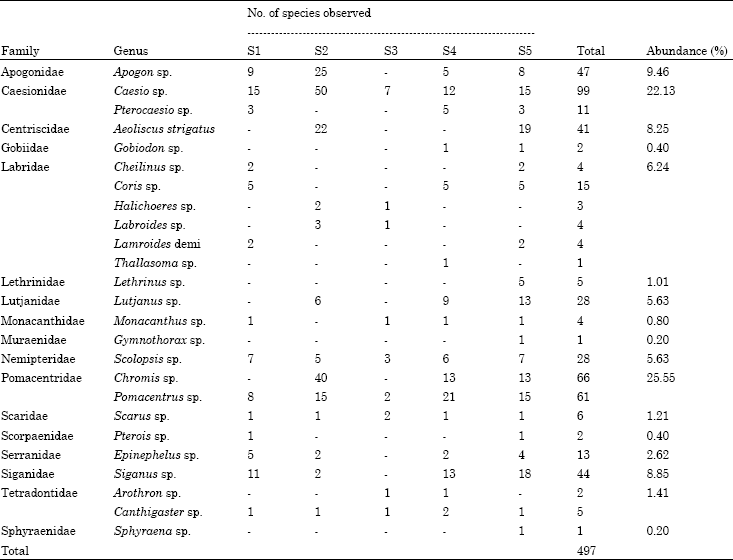
Fish distribution in the artificial seagrass area: Four hundred ninty individuals (25 genus) from 17 families were observed in 5 sampling periods throughout the study as shown in (Table 1). The identification of fishes in the artificial seagrass area was studied up to the genus level. The dominating family in the current study was Pomacentridae which occupied 25.55% of the whole area; whereas the second abundant family in the artificial seagrass area was Caesionidae which was 22.13%. Apogonidae fishes were the third dominant with representatives of Apogon sp. (9.64%) followed with Siganidae family (8.85%) and Centriscidae (8.25%). Three families were observed to be least dominant (Labridae, Lutjanidae and Nemeptiridae with 6.24, 5.63 and 5.63%, respectively) (Fig. 2). Perhaps this might indicate the artificial seagrass bed could play a significant role in attracting specific families of fishes such as Pomacentridae and Caesionidae for conservation purposes in the study area.
The dominant fish in the artificial seagrass area was Caesio sp. (family: Caesionidae) represented with 19.92% followed with Chromis sp. (Family: Pomacentridae) represented with 13.28% of the total percentage of fish observed in the study area. Thallasoma sp. (Family: Labridae), Gymnothorax sp. (Family: Muraenidae) and Sphyraena sp. (Family: Sphyraenidae) were less dominant fishes in the artificial seagrass area, each contributed with 0.2% of the total population. Shannon-Weiner index was applied to know the diversity index of fish in the artificial searass bed. The result showed that the study area quiet diverse with fish species. Out of 25 species the first 3 dominant species were Casio sp. (H’ = 0.32) followed by Chromis sp. (H’ = 0.28) and Pomacentrus sp. (H’ = 0.23).
| Table 1: | Percentage of individual fish abundance in artificial seagrass bed. (S1 - S5 = Sampling 1 to sampling 5) |
 | |
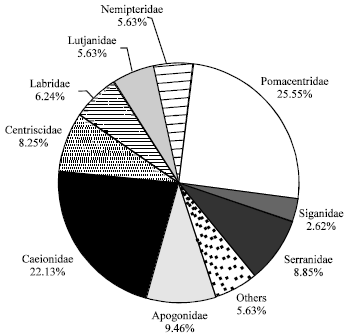 | |
| Fig. 2: | Composition of the abundance of different fish families inhabited in artificial seagrass ecosystem |
| Table 2: | Total number of species observed during the sampling period and individual species Shannon Weaver diversity Index (H’) is represented |
 | |
The least dominant species were Thallasoma sp. Gymnothorax sp. and Sphyraena sp. with H’ value of 0.01 (Table 2). The species diversity (H’) in the sampling area was 2.5315 clearly indicates that artificial seagrass bed could provide unique shelter habitat for marine fish communities.
DISCUSSION
Habitat selection by organisms is influenced by several factors including food preference, habitat structure, chemical defense, predation and competition (Hacker and Steneck, 1990; Duffy and Hay, 1991). Among this, seagrass beds are among the most widespread and productive coastal ecosystem types worldwide and range from the tropics to boreal margins of every ocean (Hemminga and Duarte, 2000). Similar findings were observed in our study area which comprised highly diversified fishes. They provide physical structure on otherwise largely featureless sediment bottoms, enhancing community diversity, biomass, primary and secondary production. Seagrass community provide nursery areas for juvenile stages of commercially important species (Heck et al., 2003) contributes significantly to the economic importance of estuarine fisheries (Anderson, 1989; Costanza et al., 1997). As such, the observed high abundance of Caesio sp. in the artificial seagrass area might be due to their feeding habit. They feed primarily on benthic invertebrates and small fishes and grow up to 18 cm in size. Due to the abundance of aquatic invertebrates like bivalves and gastropods in artificial seagrass bed might help in the feeding habit of Caesio sp. Meanwhile Chromis sp. which primarily feeds on zooplankton at offshore reefs can grow up to 8 cm (Fletcher and Fletcher, 1995). Caesio sp. and Chromis sp. those were found in the artificial seagrass area was fairly big in size (6 cm). This showed that artificial seagrass might be a suitable habitat even then the artificial seagrass area provides habitat, food and hiding place from predatory fishes (Borowitzka and Lethbridge, 1989). The genus of Thallasoma sp. (family: Labridae) Gymnothorax sp. (family: Muraenidae) and Sphyraena sp. (family Sphyraenidae) were least dominant fishes in the artificial seagrass area and each contributed only about 0.2 % to the total population. Similar finding was observed by Judy Upstona et al. (2003). He suggested that the least abundance of species belong to the families muraenidae and sphyraenidae in the artificial seagrass area proved that this habitat is not a conducive environment for their best survival. Even though some of the Muraenidae fishes can grow up to 1m and above as like Sphyraenidae fishes, however we could not generate any record proving their abundance in seagrass areas. These fishes generally prefer nearby coral reef ecosystem rather than seagrass beds. Hence we predict that artificial seagrass might not be as suitable as coral reef ecosystem for their better survival. It was also observed that they were found to be in the benthic realm shows their sluggish behavior around the artificial seagrass area.
The resulted Shannon-Weiner Diversity Index indicated that Caesio sp. (H’ = 0.32) was the most dominant fish followed by Chromis sp. (H’ = 0.29) and Pomacentrus sp. (H’ = 0.23) in the artificial seagrass area. This might be due to floating seagrass blades expedite the growth of algae as food source for these fishes and helps in providing the suitable habitat for their survival. Overall Shannon diversity value of 2.5315 clearly demonstrated that artificial seagrass bed in Sepanggar Bay provide better habitat for various species of marine fishes.
The diverse faunal communities of seagrass beds throughout South East Asia are threatened by continued overexploitation of living marine resources such as fish (Kuriandewa et al., 2003). Fish movements are utilized by local fishers across South East Asia who use Fyke nets (fish fences) set into tidal currents or between seagrass beds and reefs to catch fish. It is important for the future sustainable management of seagrass beds since it offers alternative natural habitat to sustain the marine fish communities (Parrish, 1989; Fortes, 1990).
CONCLUSION
The fish diversity in the artificial seagrass area was quite diverse based on the Shannon-Weiner Diversity Index. There were few species dominating the artificial seagrass area, which were Caesio sp., Chromis sp. and Pomacentrus sp. These fishes generally planktivorous feed primarily on phytoplankton and zooplankton, especially on the algae that grows on the artificial seagrass blades. Artificial seagrass blades can be applied to increase the abundance and the numbers of fish in a specific area. Because these structures might provide food for small fishes as algae can grow in these artificial grasses. Nevertheless the study has shown that the artificial seagrass can be used as a unique artificial habitat for marine fishes. Thus it can be utilized for the conservation of some important valued commercial fishes for the sustainable development of marine fishery resources.
ACKNOWLEDGMENTS
The authors are grateful to Dorken Reef Corporation Sdn. Bhd. No. 33B, Jalan SG 3/19, Pusat Bandar Taman Sri Gombak, 68100 Batu Caves, Selangor for supplying artificial sea grass during this research. The authors also wish to extend their gratitude to Ismail Tajul, Science officer of Borneo Marine Research Institute for species identification.
REFERENCES
- Adams, A.J., J.V. Locascio and B.D. Robbins, 2004. Microhabitat use by a post-settlement stage estuarine fish: Evidence from relative abundance and predation among habitats. J. Exp. Mar. Biol. Ecol., 299: 17-33.
CrossRef - Anderson, E.E., 1989. Economic benefits of habitat restoration: Seagrass and the Virginia hard-shell blue crab fishery. North Am. J. Fish. Manage., 9: 140-149.
CrossRefDirect Link - Barber, W.E., J.G. Greenwood and P. Crocos, 1979. Artificial seagrass-a new technique for sampling the community. Hydrobiologia, 65: 135-140.
CrossRef - Beck, M.W., K.L. Heck Jr., K.W. Able, D.L. Childers and D.B. Eggleston et al., 2001. The identification, conservation and management of estuarine and marine nurseries for fish and invertebrates. BioScience, 51: 633-642.
CrossRefDirect Link - Bell, J.D., A.S. Steffe and M. Westoby, 1985. Artificial seagrass: How useful is it for field experiments on fish and macroinvertebrates?. J. Exp. Mar. Biol. Ecol., 90: 171-177.
CrossRef - Borowitzka, M.A. and R.C. Lethbridge, 1989. Seagrass Epiphytes. In: Biology of Seagrasses: A Treatise on the Biology of Seagrasses with Special Reference to the Australian Region. Larkum, A.W.D., A.J. McComb and S.A. Shepherd (Eds.). Elsevier, Amsterdam, The Netherlands, p: 458-499.
Direct Link - Costanza, R., R. d'Arge, R. de Groot, S. Farber and M. Grasso et al., 1997. The value of the world's ecosystem services and natural capital. Nature, 387: 253-260.
CrossRefDirect Link - De Troch, M., F. Fiers and M. Vincx, 2003. Niche segregation and habitat specialisation of harpacticoid copepods in a tropical seagrass bed. Mar. Biol., 142: 345-355.
CrossRef - Duffy, J.E. and M.E. Hay, 1991. Food and shelter as determinants of food choice by an herbivorous marine amphipod. Ecology, 72: 1286-1298.
Direct Link - Duffy, J.E., 2006. Biodiversity and functioning of seagrass ecosystems. Mar. Ecol. Prog. Ser., 311: 233-250.
Direct Link - Fletcher, S.W. and W.W. Fletcher, 1995. Factors affecting changes in seagrass distribution and diversity pattern in the Indian River Lagoon Complex Between 1940 and 1992. Bull. Mar. Sci., 57: 49-58.
Direct Link - Grenouillet, G.L., D. Pont and K.L. Seip, 2002. Abundance and species richness as a function of food resources and vegetation structure: Juvenile fish assemblages in rivers. Ecography, 25: 641-650.
Direct Link - Hacker, S.D. and R.S. Steneck, 1990. Habitat architecture and the abundance and body-size dependent habitat selection of a phytal amphipod. Ecology, 71: 2269-2285.
Direct Link - Heck, K.L., C. Hays and R.J. Orth, 2003. A critical evaluation of the nursery role hypothesis for seagrass meadows. Mar. Ecol. Prog. Ser., 253: 123-136.
Direct Link - Heck, K.L. Jr. and R.J. Orth, 1980. Structural components of eelgrass (Zostera marina) meadows in the lower chesapeake bay: Decapod crustacea. Estuaries, 3: 289-295.
Direct Link - Upston, J. and D.J. Booth, 2003. Settlement and density of juvenile fish assemblages in natural, Zostera capriconi (zosteraceae) and artificial seagrass beds. Environ. Biol. Fish., 66: 91-97.
CrossRef - Khalaf, M.A. and M. Kochzius, 2002. Changes in trophic community structure of shore fishes at an industrial site in the Gulf of Aqaba, Red Sea. Mar. Ecol. Prog. Ser., 239: 287-299.
Direct Link - Lee, S.Y., C.W. Fong and R.S.S. Wu, 2001. The effects of seagrass (Zostera japonica) canopy structure on associated fauna: A study using artificial seagrass units and sampling of natural beds. J. Exp. Mar. Biol. Ecol., 259: 23-50.
CrossRefDirect Link - Nakamura, Y., M. Horinouchi, T. Nakai and M. Sano, 2003. Food habits of fishes in a seagrass bed on a fringing coral reef at Iriomote Island, Southern Japan. Ichthyol. Res., 50: 15-22.
CrossRefDirect Link - Nakamura, Y. and M. Sano, 2005. Comparison of invertebrate abundance in a Seagrass bed and adjacent Coral and sand areas at Amitori Bay, Iriomote Island, Japan. Fish. Sci., 71: 543-550.
CrossRefDirect Link - Parrish, J.D., 1989. Fish communities of interacting shallow-water habitats in tropical oceanic regions. Mar. Ecol. Prog. Ser., 58: 143-160.
Direct Link - Shannon, C.E. and W. Weaver, 1949. The Mathematical Theory of Communication. 1st Edn., University of Illinois Press, Urbana, IL., ISBN-10: 0252725484, Pages: 117.
Direct Link - Sogard, S.M. and K.W. Able, 1994. Diel variation in immigration of fishes and decopod crustaceans to artificial Seagrass Habitat. Estuaries, 17: 622-630.
CrossRef - Weinstein, M.P. and K.L. Heck, 1977. Ichthyofauna of seagrass meadows along the Caribbean coast of Panama and in the Gulf of Mexico: Composition, structure and community ecology. Mar. Biol., 50: 97-107.
CrossRefDirect Link