
J.I. Agboola
Department of Fisheries, Lagos State University, P.M.B 0001 LASU, Ojo, Lagos, Nigeria
M.A. Adewolu
Department of Fisheries, Lagos State University, P.M.B 0001 LASU, Ojo, Lagos, Nigeria
E.O. Lawson
Department of Fisheries, Lagos State University, P.M.B 0001 LASU, Ojo, Lagos, Nigeria
Journal of Fisheries and Aquatic Science
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 225-235







ABSTRACT
The Millennium Ecosystem Assessment (MA) has identified climate change as one of the main drivers of change in aquatic biodiversity and ecosystem with a very rapid increase of the impact. Recently, the Intergovernmental Panel on Climate Change (IPCC) concluded that climate change is, without doubt, occurring and that the Earth is warming. There are some climate change winners, but scientists predict that the overwhelming effect of climate change upon biodiversity including fisheries will be damaging. The nexus between climate change, fisheries and phytoplankton is particularly strong. This study considers this ‘non-fishing effects’ on fish abundance and distribution by exploring the various roles of phytoplankton in climate change and fisheries productivity. It then discusses the need to improve our knowledge of phytoplankton as mechanisms linking climate to ecosystem changes such as fisheries productivity.
PDF Abstract XML References Citation
Received: July 06, 2010;
Accepted: August 11, 2010;
Published: November 16, 2010
How to cite this article
J.I. Agboola, M.A. Adewolu and E.O. Lawson, 2011. Linking Climate Change and Fisheries: The Role of Phytoplankton. Journal of Fisheries and Aquatic Science, 6: 225-235.
DOI: 10.3923/jfas.2011.225.235
URL: https://scialert.net/abstract/?doi=jfas.2011.225.235
DOI: 10.3923/jfas.2011.225.235
URL: https://scialert.net/abstract/?doi=jfas.2011.225.235
INTRODUCTION
Climate change has been identified as one of the main drivers of change in aquatic biodiversity and ecosystem with a very rapid increase of the impact (Duraiappah, 2005). The study of the Intergovernmental panel on climate change has also made us all aware that climate change is likely to be the main driver of biodiversity loss in the future. Biodiversity (including fisheries) has already been affected by recent climate change and projected climate change for the 21st century is expected to affect all aspects of biodiversity (IPCC, 2001). It is only of recent that the Intergovernmental Panel on Climate Change (IPCC) concluded that climate change is, without doubt, occurring and that the earth is warming (IPCC, 2007). The use of plankton to reveal the impact of climate is also becoming increasingly evident as time-series lengthen (some have now been running for >50 years) and data-analysis techniques improve (Hays et al., 2005).
Studies of the global impact of climate change on wildlife have rightly made headlines as quest for understanding aquatic resource productivity and the role of ocean in carbon sequestration among other issues have provoked numerous interdisciplinary researches within the past three decades. Also, several global environmental assessments indicate that the aquatic ecosystems have been profoundly influenced by human activities (Agboola and Braimoh, 2009).
Two recently initiated international biogeochemical programmes, the Surface Ocean Lower Atmosphere Study (SOLAS, http://www.uea.ac.uk/env/solas/) (Broadgate, 2004) and Integrated Marine Biogeochemistry Ecosystem Research (IMBER, http://www.imber.info/) (Hall, 2004), co-sponsored by the International Geosphere Biosphere Programme (IGBP, http://www.igbp.kva.se/) and the Scientific Committee on Oceanic Research (SCOR, http://www.jhu.edu/wscor/) aim to improve our understanding of the consequences of environmental changes, such as increasing anthropogenic CO2. These programmes incorporates large spatial- and temporal-scale monitoring of changes in surface water CO2 and plankton activity, as well as experiments where large enclosures or ‘mesocosms’ of plankton communities are subjected to differing CO2 levels (Riebesell, 2004). In a separate initiative, the Royal Society of the UK (http://www.royalsoc.ac.uk/) has launched a review of the consequences of increased acidity levels on marine life.
Furthermore, considerable number of authors has speculated on the possible effects of climate change on fish stocks in various parts of the world (Bakun, 1990; Kennedy, 1990; Bhaud et al., 1995; Snyder et al., 2003). As one method of measuring climate change is to observe the dynamics of species that could be affected, changes in the dynamics of key commercial fishes that indicate changes in their environment occurred in the early 1990s, and possibly around 1998 (McFarlane et al., 2000). Other studies have described decadal-scale changes in North Pacific climate-ocean conditions. For example, these climate-ocean changes were associated with changes in the abundance and ocean survival of salmon (Oncorhynchus sp.), distribution and spawning behaviour of hake (Merluccius productus) and sardines (Sardinops sagax) and in recruitment patterns of several groundfish species (McFarlane et al., 2000). As a result, there is the need for improved understanding of phytoplankton (grasses of the sea)’s role as moderators of atmosphere and as the base of the oceanic food web.
The role of terrestrial and marine plants in regulating the global carbon budget is not been fully understood, nor do we know how much additional CO2 the upper ocean layers will dissolve and store if CO2 increases as predicted. These uncertainties will affect the time required for a doubling of carbon dioxide, assuming no attempt is made to reduce anthropogenic inputs to the atmosphere. The rise in global average temperature may range from 2 to 6°C (Schneider, 1989). The rate of climate change will be much faster than is thought to have occurred after the ice ages of the paleozoic eras. We do not thoroughly understand how large scale ecologic systems respond to so rapid a change, but it is instructive to attempt to forecast some of the changes and model the expected impact on ecosystems. Here, the possible role of phytoplankton is reviewed.
The need to provide unique information on the potential fate of primary production in marine ecosystems which have major effects on benthic and pelagic food webs are an essential condition for the great fisheries of world seas. To achieve this, basic researches to improve our understanding of phytoplankton as an adaptive mechanism/strategy in the face of global environmental change uncertainties will be required if sustainable fishery is to become a ‘development paradigm’.
This study briefly reviews various processes through which climate influences fisheries productivity. It then discusses the need to improve our knowledge of phytoplankton as mechanisms linking climate to ecosystem changes such as fisheries productivity. Finally, based on our present understanding we comment on ecosystem responses to future climate change using phytoplankton as proxy.
AN OVERVIEW OF PHYTOPLANKTON AND ITS GLOBAL IMPORTANCE
The word ‘plankton’ derives from the Greek ‘planktos’ meaning to ‘drift’ or ‘wander’ and is used to describe passively drifting small plants (phytoplankton) and animals (zooplankton) in aquatic systems.
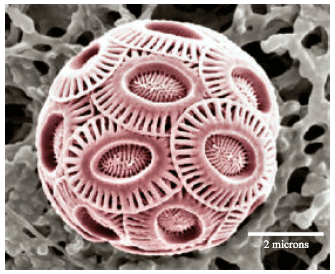 | |
| Fig. 1: | A specie of phytoplankton crucial in global carbon cycle the coccolithophore Emiliania huxleyi (Source: celsias.com) |
For example, the coccolithophore Emiliania huxleyi (Fig. 1) is widespread in oceans worldwide. This species of phytoplankton can attain massive densities (up to 20 000 cells mL-1) and cover 1000s of km2. Emiliania huxleyi is thought to produce more calcium carbonate than any other organism on Earth and, as such, has a crucial role in the global carbon cycle.
Phytoplankton contains the pigment chlorophyll, which gives them their greenish color. Chlorophyll is used by plants for photosynthesis, in which sunlight is used as an energy source to fuse water molecules and carbon dioxide into carbohydrates-plant food. Phytoplankton (and land plants) uses carbohydrates as building blocks to grow; fish and humans consume plants to get these same carbohydrates. As a result, the larger the world's phytoplankton population, the more carbon dioxide gets pulled from the atmosphere, hence, the lower the average temperature due to lower volumes of this greenhouse gas. This could also be buttressed by the fact that photosynthesis does not occur at night, leaving the available dissolved oxygen in water column utilized for respiration at night and increasing the water column dissolved carbon dioxide in early hours of the day (Agboola et al., 2008). In other words, phytoplankton responds very rapidly to changes in their environment. Large populations of this organism, sustained over long periods of time, could significantly lower atmospheric carbon dioxide levels and, in turn, lower average temperatures. As the major primary producers in the marine pelagic ecosystem, they are responsible for the transformation of dissolved inorganic carbon and nitrogen into particles. This particle formation is important for biogeochemical and ecological studies, because these biological particles can transfer to higher trophic levels unlike their dissolved counterparts because they form one part of the base of the food web. Therefore, all trophic relations and the productivity of an ecosystem are related to the primary production of phytoplankton. However, it is not the absolute primary production alone that determines the biogeochemical or ecological effect the size of the producing phytoplankton organism is one additional crucial factor. Phytoplankton size structure influences growth rate, carbon fixing capacity, nutrient uptake rate, carbon export, sedimentation rate, and the link to the food web which are supposed to be higher in areas of high production with a community dominated by large phytoplankters. Eutrophic areas of the ocean are generally dominated by larger phytoplankton like diatoms. Their cell size can reach to several hundred-ìm in diameters.
During the last two decades it has been established that cell size distribution of primary producers plays a significant role within the community structure and in the trophic transfer of the pelagic ecosystem (Malone, 1980; Azam et al., 1983). Most of the studies have shown that an important fraction of the autotrophic biomass in different regions consists of small cell size algae populations (<10 μm), which may account for 40-75% of the total primary production and 35-60% of the total phytoplankton biomass (Tremblay and Legendre, 1994). In highly productive upwelling ecosystems, the abundance of large (>10 μm) phytoplankton often leads to high values of primary production, new (or nitrate based) production, and carbon export (Michaels and Silver, 1988; Owens et al., 1991). We know today that picoplankton is a major component in all warm-water marine ecosystems, and their discovery changed our perception of marine food webs substantially. As a consequence, size is becoming an important ecological variable of the structure and functioning of food webs by having strong influence on efficiency of transfer and the fate of carbon to higher trophic levels (Michaels and Silver, 1988; Legendre and Le Fevre, 1989). Moreso, phytoplankton community structure can determine biomass, productivity, and carbon export in marine systems, thus, the need for this important review and the need for further research in elucidating the drivers of phytoplankton size and community structure.
Climatic trend: Recent observations show that greenhouse gas emissions and many aspects of the climate are changing near the upper boundary of the IPCC range of projections. Many key climate indicators are already moving beyond the patterns of natural variability within which contemporary society and economy have developed and thrived. These indicators include global mean surface temperature, sea-level rise, global ocean temperature, Arctic sea ice extent, ocean acidification, and extreme climatic events. With unabated emissions, many trends in climate will likely accelerate, leading to an increasing risk of abrupt or irreversible climatic shifts.
Since, the last IPCC report, updated trends in surface ocean temperature and heat content have been published (Domingues et al., 2008; Church et al., 2009). These revised estimates show (Fig. 2) that the ocean has warmed significantly in recent years. Current estimates indicate that ocean warming is about 50% greater than had been previously reported by IPCC (2007). The new estimates help to better explain the trend in sea level that has been observed in recent decades as most of the sea-level rise observed until recently has been the result of thermal expansion of seawater.
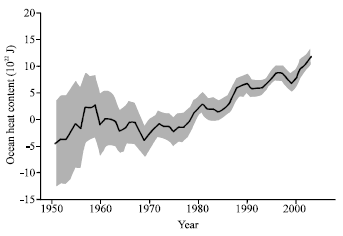 | |
| Fig. 2: | Change in ocean heat content since 1951 (observations-black line) with uncertainties (in grey shading), relative to the ocean heat content in 1961(Domingues et al., 2008) |
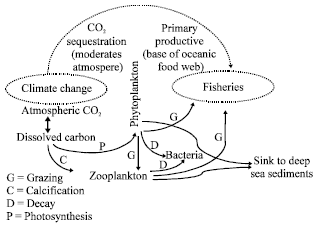 | |
| Fig. 3: | Simplified interaction of phytoplankton to climate change and fisheries |
The nexus of climate change, fisheries and phytoplankton: The nexus between climate change, fisheries and phytoplankton is particularly strong. Phytoplankton and terrestrial plants fix atmospheric CO2 to produce carbohydrates that fish and humans consume. As for fisheries, the impacts of climate change on aquatic systems are already apparent in many parts of the world, with accelerating impacts likely for several decades irrespective of future.
Phytoplankton is the foundation of marine food chain and they can influence earth’s climate. A simplified interaction to climate change and fisheries is presented in Fig. 3. Like terrestrial plants, they require sunlight, water, and nutrients for growth. Because sunlight is most abundant at and near the sea surface, phytoplankton remains at or near the surface. They are responsible for about half the photosynthesis on earth. They convert light and nutrients into organic material which is then fed right the way up through the food chain, making them key elements of life on the planet (Behrenfeld et al., 2006). After dying, some of the plants drift down into the deep ocean and lock up carbon for very longer periods. The tiny plants live in the top layer of the ocean, but rely on nutrients that are brought up from deeper in the ocean. This mixing relies on convective currents being able to break through the ‘stratification’ of water. This layering arises because layers of water at different temperatures have different densities and therefore do not mix. If the top layer of ocean water warms, as it has done, the density contrast between it and the layer below becomes greater. This makes it harder for the upwelling nutrients to reach the surface, starving the phytoplankton. Some researchers found that drops in the amount of chlorophyll being detected by the satellite closely corresponded to increases in surface water temperature, confirming the predictions of climate models (Hays et al., 2005).
Now that the concentration of carbon dioxide in atmosphere is constantly increasing due to anthropogenic emissions by about 1 ppm year-1 (Keeling and Bacastow, 1977) and is presently around more than 30% higher than before the industrial revolution (Keeling and Whorf, 2000), the need for a proactive approach to sustaining our aquatic systems through better institutionalized framework is more felt than ever.
In aquatic systems, nutrient availability sets limits on production; changes in upwelling, increased stratification, or changes in runoff patterns could reduce or shift nutrient inputs and cause swings in productivity. Agboola et al. (2010) reported the impact of nearby presence of ports on adjacent coastal waters, leading to alteration in nutritional environment and hence affecting phytoplankton biomass build-up. Also, there are situations where nutrient-rich oceanic waters are limited by deficiencies in trace metals such as iron (Martin and Fitzwater, 1988). Such, deficiencies may affect carbon dioxide fluxes from ocean to atmosphere. Conversely, production may be stimulated by windborne deposition of terrestrial trace metals; thus, with climate change, greater fluctuations in oceanic productivity will affect carbon exchange and ultimately fish populations.
CLIMATE CHANGE, FISHERIES AND THE UNCERTAINTIES
It is now widely accepted that the earth’s climate is changing under the influence of anthropogenic activities. A number of key changes in the earth’s atmosphere and ocean have already been detected (including increasing global surface temperature, rising sea levels, increases in incident UV radiation, changes in average annual precipitation, and increases in the variability and intensity of extreme weather events, among others), while speculation regarding future changes is rife (Clark, 2006). Adewolu et al. (2009) stressed on the environmental issues that have to be addressed if fisheries (aquaculture) must realize its full potential in Sub-Saharan Africa. Such environmental issues include aquatic pollution as a result of urbanization and industrialization, sand filling and dredging of harbor, pesticide run-off from agricultural land, conflict of land use between aquaculture and industries, tourism and climate change effects.
In relation to Fig. 4, a trophic level of an organism is its position in a food chain. Levels are numbered according to how far particular organisms are along the chain from the primary producers at level 1, to herbivores (level 2), to predators (level 3), and to carnivores or top carnivores (level 4 or 5). Fish at higher trophic levels are typically of higher economic value. The decline in the trophic level harvested is largely a result of the over-harvest of fish at higher trophic levels. A decrease in plankton production, which could result from a changing climate, would mean less food for fish populations. Globally, the degradation of fisheries is also reflected in the fact that the fish being harvested are increasingly coming from the less valuable lower trophic levels as populations of higher trophic level species are depleted.
The current impact and trend of climate change and other drivers on aquatic biodiversity and ecosystem is shown on Table 1. Climate change impacts are highlighted as low to moderate over the last century and current impact as very rapid when compared with other drivers, with expected increasing importance over the next 50 years. This importance is tightly linked with the ability to keep temperatures below a 2°C increase (Duraiappah, 2005).
Increased atmospheric temperatures which is perceived to result in changes in local weather patterns; hydrology regimes, changes to the hydrology cycle, changes in runoff and river discharge, and changes in timing of hydrologic events; changes in coastal currents, in sea surface temperatures, in ocean-atmosphere interactions, in open ocean circulation patterns and changes in upwelling systems that might take place with climatic change.
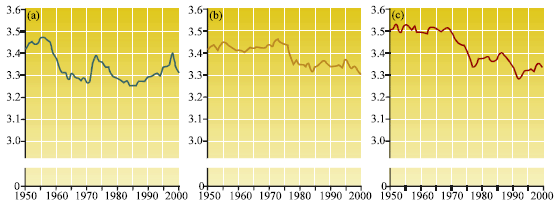 | |
| Fig. 4: | Decline in trophic level of fisheries catch since 1950 (MEA, 2005) |
| Table 1: | Current impact and trends of climate change and other drivers on aquatic biodiversity (Fisheries) and ecosystems Adapted from (MEA, 2005) |
 | |
▲: Low: ▲▲: Moderate, ▲▲▲: High, ▲▲▲▲: Very high. x: Continuing impact, xx: Increasing impact, xxx: Very rapid increase of the impact | |
Fish populations will respond to climatic change with an expected impact on marine primary productivity and on trophic regimes. In addition, temperature increases are expected to shift many species polewards where the climate is cooler and hence metabolic rates could be kept low. Warm-water species may increase in abundance with more favourable conditions at lower latitudes. This situation could have a significant impact on commercial fisheries such as that of Atlantic cod as this species feeds on temperature-sensitive ones like mackerel and herring. Research into the decline of important salmon stocks on the west coast of Canada and Alaska has suggested that these temperature changes are already having an impact. Natural climate variability experienced during the El Niño Southern Oscillation (ENSO) event provides further warning of the potential impacts of climate change. In the past ENSO events have led to a mass die-off of fish species such as Peruvian anchovies and sardines (Hays et al., 2005). Further large-scale mortalities could result if the unusual temperatures experienced during an ENSO period become more commonplace with climate change (Hughes, 2000).
The eggs and larvae of many fish species are sensitive to ultraviolet (UV) radiation (Gucinski et al., 1990). The naturally high mortality of fish larvae make it difficult to accurately assess the direct effects of ozone depletion when the primary causes of population decline are predation, poor food supply for larvae, overfishing of adults, water temperature, pollution and disease. Nonetheless, UV radiation appears to be harmful at both primary and secondary production levels. The number of variables involved makes predictions of impacts on a global scale and further along the food web a complex task.
Different global climate models have significantly different projections for the local effects of climate change. The exact relationship between atmospheric CO2 and global temperature rise is also under debate and we do not know the scale of greenhouse emissions over the coming decades (Behrenfeld et al., 2006). The scientific models that assess the impact of climate change on biodiversity are built upon these uncertainties, and are relatively simplistic in comparison to the multitude of factors that will affect species distributions in the future. The links between biodiversity and climate change run both ways: biodiversity is threatened by climate change, but proper management of biodiversity can reduce the impacts of climate change. Climate fluctuations in North America reduce plankton populations, the main source of food of the North Atlantic right whale. Only about 300 individuals remain at present and the reduced availability of food due to climate change is becoming an increasing cause of mortality (Gucinski et al., 1990).
The role of phytoplankton in carbon sequestration and productive fisheries: Scientists have found that a given population of phytoplankton can double its number on the order of once per day. In other words, phytoplankton responds very rapidly to changes in their environment. Changes in the trends for a given phytoplankton population such as its density, areal distribution, and rate of population growth or diminution-should alert Earth scientists of changes in environmental conditions. Then, by comparing these phytoplankton trends to other measurements such as temperature scientists can learn more about how phytoplankton may be contributing to, and affected by, climatic and environmental change. Thus, phytoplanktons are indicators of change. They are responsible for removing carbon dioxide from the atmosphere and transferring the carbon to other trophic levels. Phytoplankton also provides a vital link to deep ocean organisms when they die and organic material falls to the seabed. They are found in the top layer (the euphotic zone) where they receive enough solar radiation for their photosynthetic requirements as well as nutrients that are transported by the upwelling process. Several authors have summarized relationships between primary production and fisheries yield (Ryther, 1969; Oglesby, 1977). Nixon (1988) reviewed and re-analyzed these data and reported that fisheries yield varied as primary production was raised to the 1.55 power, with a correlation coefficient of 0.84; thus, a 5% reduction in primary productivity would lead to a 6 to 9% reduction in fisheries yield (Hardy and Gucinski, 1989).
There is strong evidence for systematic changes in plankton abundance and community structure over recent decades in many areas worldwide. Antarctic krill, which are a key component of the diet of whales and other marine mammals, have declined in abundance by more than an order of magnitude during the past 25 years (Atkinson et al., 2004). This decline has been linked to reduced food availability for krill in the form of phytoplankton blooms in summer and ice algae in winter.
Presently, our knowledge of the impacts of temperature change on phytoplankton populations is poor and reliant on predictions of changing physical processes. According to Hays et al. (2005), two such changes that could result from an increase in sea surface temperature are altered wind patterns and increased stratification of the water column. These effects could significantly reduce nutrients reaching the euphotic zone in certain parts of the world. This would lead to a decrease in primary production which would mean less overall productivity as well as less carbon dioxide removed from the atmosphere. The deep sea ecosystem would also be affected by a reduced carbon input.
The reduction in numbers or change in species composition of phytoplankton could lead to a reduction in zooplankton abundance. Since the 1950s scientists have recorded a decrease in zooplankton numbers in the Californian current. Whether this steady decline is attributable to a decrease in primary production or an increase in predation on the zooplankton, is not certain.
Alterations in nutrient levels and temperature may also effect the species composition of phytoplankton populations in different regions, which would have serious repercussions for the rest of the marine environment. Not all types are readily eaten by zooplankton, some are even toxic, and different species also absorb carbon dioxide to varying degrees. A shift in species in any one area could lead to less carbon dioxide being absorbed from the atmosphere.
Plankton is not only affected by temperature changes but the increase in ultraviolet (UV) radiation, as a result of ozone depletion, also poses a threat to their survival. Studies have shown that UV radiation actually inhibits photosynthesis in phytoplankton but it is difficult to quantify these effects in the open ocean due to variation caused by factors such as vertical mixing and cloud cover (Behrenfeld et al., 2006). It is thought that rapid vertical mixing in the water column further inhibits photosynthesis whereas a greater cloud cover reduces the harmful effects of UV radiation. In addition UV rays affect growth and reproduction as well as the functioning of enzymes and cellular proteins in phytoplankton. Damage to phytoplankton has been demonstrated at the molecular, cellular, population and community levels but the effects of increased UV radiation at the ecosystem level remain uncertain.
CONCLUSION
The capacity of aquatic systems to sustain fisheries fundamentally depends on the existence and availability of primary producers, the plants that fix carbon dioxide into living organic matter (Gucinski et al., 1990). Presently, demand has developed for collaboration of scientists with public policy agencies, business, labor and NGOs to assure that research findings/solutions find their way into practical applications and policy. This paper proposes that a new research platform be geared towards identifying the factors that determine species assemblage structure of primary producers. To achieve this, there would be need to embark on a regional scales research. Results obtained from regions will enable us identify the factors that determine species assemblage structure of primary producers, which is critical to improving our understanding of controls on the strength of the biological pump and for making predictions about production at higher trophic levels. This has important implications for a range of issues from the success of commercially important fish species to marine biodiversity. Thus, the need for basic research on topics where lack of understanding prevents design of adaptive management/policy cannot be overemphasized.
To conclude in the words of Behrenfeld (New Scientist, 2006), “What was particularly astonishing to me was just how well the climate indices that we were using matched the changes of plants in water”. With this assertion, therefore, provision of adequate spatial coverage in synoptic or quasi-synoptic surveys, associated with the extreme temporal and spatial variability of the physical and biological properties especially in the dynamic coastal ecosystems will make assessment of overall phytoplankton patterns less complicated. Effort geared towards filling these information gaps may be of unique benefit to coastal ecosystem dynamic studies and policy makers on the sustainable use and management of coastal water resources.
ACKNOWLEDGMENT
We would like to thank the Lagos State University Library for providing us with some of the materials used for this review.
REFERENCES
- Adewolu, M.A., S.L. Akintola, A.A. Jimoh, F.G. Owodeinde, O.O. Whenu and K.A. Fakoya, 2009. Environmental threats to the development of aquaculture in Lagos State, Nigeria. Eur. J. Sci. Res., 34: 337-347.
Direct Link - Agboola, J.I., M. Uchimiya, I. Kudo, K. Kido and M. Osawa, 2010. Dynamics of pelagic variables in two contrasting coastal systems in the western Hokkaido coast off Otaru port, Japan. Estuarine, Coastal Shelf Sci., 86: 477-484.
CrossRef - Agboola, J.I. and A.K. Braimoh, 2009. Strategic partnership for sustainable management of aquatic resources. Water Resour. Manage., 23: 2761-2775.
CrossRef - Agboola, J.I., M.A. Anetekhai and A.A.B. Denloye, 2008. Aspects of the ecology and fishes of Badagry creek (Nigeria). J. Fish. Aquat. Sci., 3: 184-194.
CrossRefDirect Link - Atkinson, A., V. Siegel, E. Pakhomov and P. Rothery, 2004. Long-term decline in krill stock and increase in salps within the southern Ocean. Nature, 432: 100-103.
Direct Link - Azam, F., T. Fenchel, J.G. Field, J.S. Gray, L.A. Meyer-Reil and T. Thingstad, 1983. The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser., 10: 257-263.
CrossRefDirect Link - Bakun, A., 1990. Global climate change and intensification of coastal global upwelling. Science, 247: 198-201.
CrossRef - Bhaud, M., J.H. Cha, J.C. Duchene and C. Nozais, 1995. Influence of temperature on the marine fauna: What can be expected from a climatic change. J. Thermal Biol., 20: 91-104.
CrossRef - Behrenfeld, M.J., R.T. O'Malley, D.A. Siegel, C.R. McClain and J.L. Sarmiento et al., 2006. Climate-driven trends in contemporary ocean productivity. Nature, 444: 752-755.
Direct Link - Church, J., C. Domingues, N. White, S. Wijffels, P. Barker and P. Gleckler, 2009. Changes in global upper-ocean heat content over the last half century and comparison with climate models. IOP Conf. Ser.: Earth Environ. Sci., 6: 032005-032005.
Direct Link - Clark, B.M., 2006. Climate change: A looming challenge for fisheries management in southern Africa. Marine Policy, 30: 84-95.
CrossRef - Domingues, C.M., J.A. Church, N.J. White, P.J. Gleckler, S.E. Wijffels, P.M. Barker and J.R. Dunn, 2008. Improved estimates of upper-ocean warming and multi-decadal sea-level rise. Nature, 453: 1090-1093.
Direct Link - Gucinski, H., R.T. Lackey and B.C. Spence, 1990. Global climate change: Policy implications for fisheries. Fisher. Bull. Am. Fisher. Soc., 15: 33-38.
Direct Link - Hardy, J. and H. Gucinski, 1989. Stratospheric ozone depletion: Implications for marine ecosystems. Oceanography Mag., 2: 18-21.
Direct Link - Hays, G.C., A.J. Richardson and C. Robinson, 2005. Climate change and marine plankton. Trends Ecol. Evol., 20: 337-344.
CrossRef - Hughes, L., 2000. Biological consequences of global warming: Is the signal already apparent. Trends Ecol. Evol., 15: 56-61.
CrossRefDirect Link - IPCC, 2007. Climate Change 2007: The Physical Science Basis. In: Contribution of Working Group I to the 4th Assessment Report of the International Panel on Climate Change Solomon, Solomon, S., D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor and H.L. Miller (Eds.). Cambridge University Press, Cambridge, New York, USA., ISBN: 978052188009-1, pp: 996-996.
Direct Link - Kennedy, V.S., 1990. Anticipated effects of climate change on estuarine and coastal fisheries. Fisheries, 15: 16-24.
Direct Link - Martin, J.H. and S.E. Fitzwater, 1988. Iron deficiency limits phytoplankton growth in the north-east pacific subarctic. Nature, 331: 341-343.
Direct Link - McFarlane, G.A., J.R. King and R.J. Beamish, 2000. Have there been recent changes in climate? Ask the fish. Prog. Oceanography, 47: 147-169.
Direct Link - Nixon, S.W., 1988. Physical energy inputs and the comparative ecology of lakes and marine ecosystems. Limnol. Oceangr., 33: 1005-1025.
Direct Link - Oglesby, R.T., 1977. Relationships of fish yield to lake phytoplankton standing crop, production and morphoedaphic factors. J. Fish. Res. Board Can., 34: 2271-2279.
Direct Link - Owens, N.J.P., J. Priddle and M.J. Whitehouse, 1991. Variations in phytoplankton nitrogen assimilation around South Georgia in the Bransfield strait (Southern Ocean). Mar. Chem., 35: 287-304.
CrossRef - Riebesell, U., 2004. Effects of CO2 enrichment on marine phytoplankton. J. Oceanogr., 60: 719-729.
Direct Link - Snyder, M.A., L.C. Sloan, N.S. Diffenbaugh and J.L. Bell, 2003. Future climate change and upwelling in the California current. Geophys. Res. Lett., 30: 1823-1824.
Direct Link - Tremblay, J.E. and L. Legendre, 1994. A model for the size fractionated biomass and production of marine phytoplankton. Limnol. Oceanography, 39: 2004-2014.
Direct Link