
E. Espino-Barr
CRIP-Manzanillo, INAPESCA, Playa Ventanas s/n, Manzanillo, Colima, 28200, Mexico
M. Gallardo-Cabello
Unidad de Ecolog�a Marina, Instituto de Ciencias del Mar y Limnolog�a, Universidad Nacional Aut�noma de Mexico, D.F.C.P. 04510, Mexico
K. Granados-Flores
Unidad de Ecolog�a Marina, Instituto de Ciencias del Mar y Limnolog�a, Universidad Nacional Aut�noma de Mexico, D.F.C.P. 04510, Mexico
E.G. Cabral-Solis
CRIP-Manzanillo, INAPESCA, Playa Ventanas s/n, Manzanillo, Colima, 28200, Mexico
A. Garcia-Boa
CRIP-Manzanillo, INAPESCA, Playa Ventanas s/n, Manzanillo, Colima, 28200, Mexico
M. Puente-Gomez
CRIP-Manzanillo, INAPESCA, Playa Ventanas s/n, Manzanillo, Colima, 28200, Mexico
Journal of Fisheries and Aquatic Science
Year: 2010 | Volume: 5 | Issue: 4 | Page No.: 293-303







ABSTRACT
Thirteen age groups were identified for Microlepidotus brevipinnis (Pisces: Haemulidae). Otoliths sagittae gave a better fit between observed and calculated data than scales, but both methods are considered valid for age determination. The constants of Von Bertalanffy’s growth equation calculated through sagittae analysis were: L∞ = 388.68 mm, W∞ = 829.741 g, K = 0.207, to = -0.718 and A0.95 = 14 years. Differences in growth between sexes were found; females show higher values of L∞ and longevity than males. Microlepidotus brevipinnis reaches its sexual maturity at age four. The gonadosomatic index presented higher values during December and March. The periods of more intensive feeding and fatty acids accumulation in liver are from May to October. With other researcher data, growth parameters of species from the same family were calculated in order to compare them with the one obtained in the present study. Microlepidotus brevipinnis has an intermediate size among other species of the same family.
PDF Abstract XML References Citation
How to cite this article
E. Espino-Barr, M. Gallardo-Cabello, K. Granados-Flores, E.G. Cabral-Solis, A. Garcia-Boa and M. Puente-Gomez, 2010. Growth Analysis of Microlepidotus brevipinnis from the Pacific Coast of Jalisco, Mexico. Journal of Fisheries and Aquatic Science, 5: 293-303.
DOI: 10.3923/jfas.2010.293.303
URL: https://scialert.net/abstract/?doi=jfas.2010.293.303
DOI: 10.3923/jfas.2010.293.303
URL: https://scialert.net/abstract/?doi=jfas.2010.293.303
INTRODUCTION
Microlepidotus brevipinnis (Pisces: Haemulidae) or brassy grunt (Fig. 1) is a demersal and coastal species of the continental shelf from Baja California and the Gulf of California to Chiapas, Mexico (Allen and Robertson, 1994; Fischer et al., 1995; Nelson et al., 2004). It is commonly fished during the whole year by artisanal fishers off the coast of Jalisco, México. Its capture volumes constitute about 10% of total annual average catch (Cruz-Romero et al., 1993).
 | |
| Fig. 1: | Brassy grunt Microlepidotus brevipinnis |
This species is considered a second class commercial product; its price is from 20 to 30 pesos on the beach or from 30 to 40 pesos per kg in the market ($2.15 to $2.85 USD). Analysis on its taxonomic description has been made by Castro-Aguirre et al. (1999), Van der Heiden (1985) and Chavez-Ramos et al. (1994). Its fishery has been described by Ramirez-Granados (1977), Cruz-Romero et al. (1989, 1993, 1996) and Espino-Barr et al. (2003, 2004a) analyzed age and growth of this species using and indirect method of length frequency. Granados-Flores et al. (2010) determined the average length for each age with the analysis of the sagittae and scales.
Age and growth studies are necessary to understand the behavior of the fish population in time and space (Lucano-Ramirez et al., 2006). Analysis of age through direct methods based on hard structures as otoliths and scales are good instruments to estimate age groups and obtain growth Von Bertalanffy parameters (Espino-Barr et al., 2008; Gallardo-Cabello et al., 2006).
Analysis of the weight-length relation is very important to obtain the condition factor which explains yearly weight variations related to food availability (Safran, 1992). Calculations of the gonadosomatic index and the hepatosomatic index help to understand the growth through fish life span, the gonadic maturity and spawning phenomena, the fatty acid accumulation in the liver and the trophic dynamics of the populations (Saucedo-Lozano et al., 1999; Rojas-Herrera et al., 2004; Espino-Barr et al., 2004b, 2008; Rojas et al., 2004; Aguirre-Leon and Diaz-Ruiz, 2006).
In this study, the growth constants by Von Bertalanffy’s (1938) equation were calculated from the observed data obtained by the analysis of otoliths and scales. Differences between sexes were also analyzed. Longevity and values of the condition factor, gonadosomatic and hepatosomatic indexes were calculated.
MATERIALS AND METHODS
From January 2005 to June 2006, 1050 specimens of Microlepidotus brevipinnis (Pisces: Haemulidae) were obtained from the commercial catch in Melaque, coast of Jalisco (104°40’30” to 104°43’30”W and 19°12’00” to 19°13’30”N) (Fig. 2).
Standard (Ls, cm) and total length (Lt, cm), total (Wt, g) and eviscerated weight (We, g), gonad (Wg, g) and liver weight (Wl, g) were recorded for each individual. Sex was determined for 480 individuals; the other 570 were eviscerated by fishers before landing. For age study two structures were obtained from 353 individuals: otoliths (sagittae) and scales.
The average length of the fish at age groups determined by the analysis of the otoliths sagittae and scales by Granados-Flores et al. (2010) were used to obtain the constants by Von Bertalanffy’s (1938) growth equation. The observed values determined by sagittae were: for age 1 = 125.0 mm; age 2 = 141.72 mm; age 3 = 222.32 mm; age 4 = 246.46 mm; age 5 = 283.37 mm; age 6 = 291.76 mm; age 7 = 304.09 mm; age 8 = 318.44 mm; age 9 = 335.19 mm; age 10 = 340.46 mm; age 11 = 349.54 mm; age 12 = 367.24 mm and age 13 = 372.68 mm. The observed values determined by scales were: for age 1 = 125.0 mm, age 2 = 138.80 mm; age 3 = 220.00 mm; age 4 = 243.80 mm; age 5 = 281.50 mm; age 6 = 285.60 mm; age 7 = 295.20 mm; age 8 = 307.10 mm; age 9 = 315.60 mm; age 10 = 331.10 mm, age 11 = 346.40 mm; age 12 = 354.00 mm and age 13 = 360.50 mm.
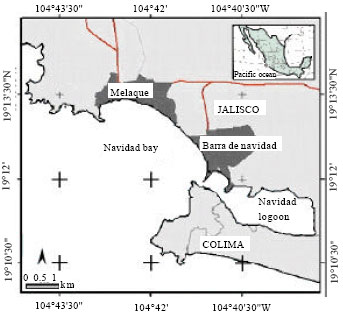 | |
| Fig. 2: | Map of the study area, San Patricio Melaque, Jalisco, Mexico |
The time of formation of slow or fast growth bands in sagittae, as well as the time of ring formation in scales was one year. The growth of otoliths and scales was proportional to the growth of the fish. There were growth differences between sexes (Granados-Flores et al., 2010).
The constants L∞, K and to of Von Bertalanffy’s (1938) equation were obtained with Ford (1933), Walford’s (1946) and Gulland’s method and were adjusted by convergent iterations of Newton’s algorithm with the Solver program in Excel software (Microsoft, 1992).
Weight-length relationship was calculated with the function W = a * L b (Mendenhall, 1987; Zar, 1996). Weight for every age was obtained with the growth data in length and the weight-length function. Weight growth was obtained substituting Lt and L∞ for Wt and W∞, in Von Bertalanffy’s (1938) equation. Age limit or longevity (95% of L∞) was determined by Taylor (1958, 1960) equation:
Monthly values of the condition factor, equivalent to the a parameter of weight-length equation, were obtained for total and eviscerated weight (Safran, 1992) and compared with the confidence interval, to explain changes in the gonad and liver throughout the year.
The gonadosomatic index (GSI) was determined with the equation described by Rodriguez-Gutierrez (1992): GSI = Wg/Wt*100, where Wg is gonad weight and Wt is the total weight of the individual.
The hepatosomatic index (HSI) was calculated by Rodriguez-Gutierrez (1992) as:
where, Wl is the liver weight and Wt is the total weight of the individual.
RESULTS AND DISCUSSION
Growth in Length
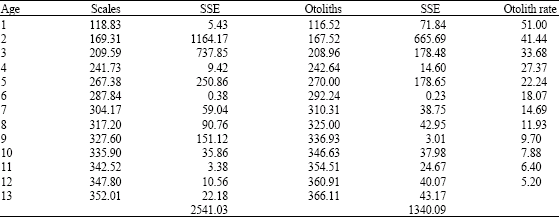
Analysis of the sagittae and scales permitted to identify thirteen age groups. Growth parameters for the total length determined through sagittae were: L∞ = 388.68 mm, K = 0.207 years-1, to = -0.718 and for scales were L∞ = 368.66 mm, K = 0.226 years-1, to = -0.724.
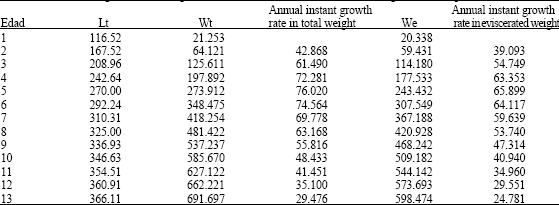
Estimated length for each age for otoliths and scales are shown in Table 1. Sum Square Error (SSE) shows that otoliths gave a better fit between observed and calculated data than scales. So, the sagittae can be considered a better structure for age determination of M. brevipinnis. Instantaneous growth rate is also shown in Table 1. During the first year of age M. brevipinnis reaches 116.52 mm of length after which growth rate decreases to 51 mm between ages 1 and 2 and 5.2 mm in ages 12 and 13 years.
Growth in Weight
The allometric growth index value of the total weight-length equation was isometric: b = 3.042. For eviscerated specimens, b = 2.954, which shows a tendency to negative allometric growth (Fig. 3a, b).
| Table 1: | Total length values for each age estimated with the Von Bertalanffy (1938) growth equation and fitted with solver program for M. brevipinnis |
 | |
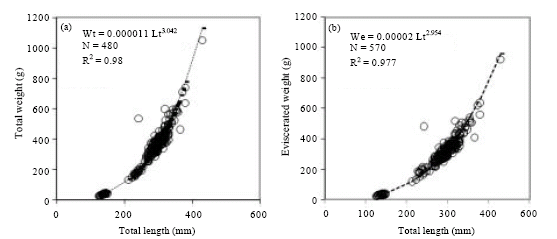 | |
| Fig. 3: | Weight (g) and length (mm) relationship of M. brevipinnis: (a) total weight and (b) eviscerated weight |
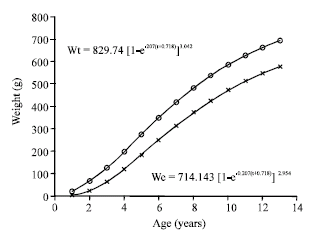 | |
| Fig. 4: | Von Bertalanffy’s growth curve in weight (g) of M. brevipinnis |
| Table 2: | Estimated length (Lt) and weight: total (Wt) and eviscerated (We) of each age of M. brevipinnis |
 | |
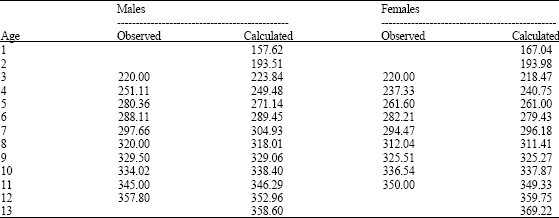
| Table 3: | Length values of otoliths for each age estimated by sexes with the von Bertalanffy (1938) growth equation for M. brevipinnis |
 | |
Theoretical growth in weight: Table 2 and Fig. 4 show the values of theoretical growth for total and eviscerated weight at each age. The asymptotic total weight was Wt∞ = 829.741 g and the eviscerated asymptotic weight, We = 714.143 g.
Differences Between Sexes
Growth parameters values from otoliths by sexes of M. brevipinnis for males were: L∞ = 381.81 mm, K = 0.197 years-1, to = -1.122 and L∞ = 489.05 mm, K = 0.082 years-1; to = -4.251 (Fig. 5a, b) for females. Table 3 shows length values of otoliths for each age by sexes.
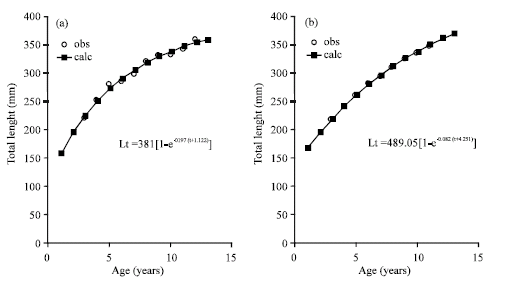 | |
| Fig. 5: | Von Bertalanffy’s growth curve in length (mm) with otoliths by sexes of M. brevipinnis: (a) males and (b) females |
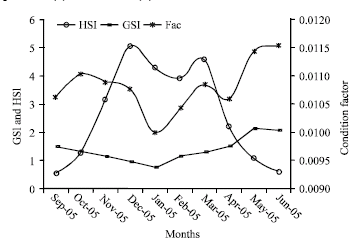 | |
| Fig. 6: | Monthly variations of the gonadosomatic (GSI) and hepatosomatic (HSI) index and condition factor in M. brevipinnis |
Longevity (Age A0.95)
Microlepidotus brevipinnis reached 95% of its infinite length L∞ at 14 years. Differences between sexes are: males 14 years and females 32 years.
Bioecologic Considerations
Figure 6 shows the trends of the monthly average data of the indexes of condition factor (a), gonadosomatic (GSI) and hepatosomatic (HSI). Massive spawning period occurs from December to March with two important pulses and two less abundant (January and February). The most active feeding season represented as condition factor (Fac) is from May to October. The hepatosomatic index (HSI) reaches its highest values in May and June and decreases abruptly in December and January.
Growth Parameters of Members of the Haemulidae Family
Table 4 shows growth parameters L8 and K of 23 species of the Haemulidae family around the world.
| Table 4: | Values of the growth parameters for different species of the Haemulidae family |
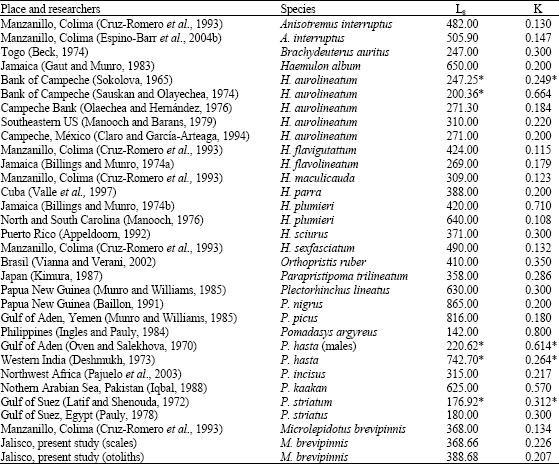 | |
*Calculated by the researchers of this study | |
In this study, the values of growth obtained by the identification of growth rings in the sagittae were very similar to those found with scales analysis which rendered age determination in M. brevipinnis valid with both methods and validating them both.
The high growth rate in length during the first year of life shows M. brevipinnis reduction of natural mortality by avoiding depredation.
In the relationship of the standard length vs eviscerated weight, Cruz-Romero et al. (1993) found an allometric index of 2.45 which represent a thinner organism. This difference between years is closely related to environmental conditions and food availability (Safran, 1992).
Microlepidotus brevipinnis reaches reproduction maturity at age 4. From age 1 to age 4 growth curves for total and eviscerated organisms are equal. Both curves separate after age 4, probably because the weight of the reproductive organs has started to mature.
Females reach a lower catabolic index than males; therefore, their asymptotic length and longevity are higher than in males. After age 4 females use most of its energy to produce reproductive products (eggs).
The three indexes: condition factor (Fac), gonadosomatic (GSI) and hepatosomatic (HSI), show that the main spawning period occurs from December to March. During this period it was observed that the condition factor starts to reduce, individuals loose weight and the HSI reaches its lowest values due to loss of fatty acid reserves.
The period during which Fac increases (May to October) coincides with the rainy season and hurricanes, when food availability is higher, due to the delivery of nutriments by the rivers to the coastal zone and the removal of the bottom to the surface of the sea due to the winds (Manzanillo’s Observatory, personal communication). This is a period in which specimens increase their individual weight, reserves of liver fatty acids and prepare for the next spawning season.
The value of L∞ of Von Bertalanffy’s (1938) growth equation of this study (368.66 mm with scales) in relation to the one found a decade ago (368.00 mm by length frequencies distribution) by Cruz-Romero et al. (1993) are almost the same suggesting certain stability in the population over the years in spite of environmental fluctuations and fishing pressure during the last 25 years; a reduction in length has not been observed, which could be considered as an overfishing symptom. Cruz-Romero et al. (1993) calculated a lower longevity (23 years) than the one found in the present study, which relates to a lower value of the catabolic index of K = 0.134.
Besides time periods, differences in the results of the study of Cruz-Romero et al. (1993) and ours can be due to the methods: they used an indirect method with size frequency analysis and we used two direct methods: scales and otoliths, which can be more accurate.
Genus Plectorhinchus reaches the largest L8 with the species P. nigrus (865 mm), P. picus (816 mm) and P. lineatus (639 mm). Other large Haemulidae species are Pomadasys hasta (742.7 mm), Haemulon album (650 mm), H. plumieri (640 mm), P. kaakan (625 mm) and Anisotremus interruptus (505.9 mm). Except for H. plumieri, most of these specimens are found in tropical areas where the temperature is higher. The smaller species of this family are the Pomadasys genus: P. argyreus (142 mm), P. striatum (176.9 mm) and P. striatus (180 mm). According to the value of L8, M. brevipinnis has a middle size with respect to other members of the Haemulidae family.
Haemulon aurolineatum is the most studied species, five papers have been reported, the ones from the Bank of Campeche have lower L8 than that of Southeastern US, which is in accordance with Taylor (1958, 1960), who established that there is a direct relation between L8 and latitude and an inverse relation between the L8 and temperature. This also occurs with H. plumieri in the areas of Jamaica and Carolina. In this case, it can also be observed that the catabolic index K is lower when the latitude increases and temperature decreases.
Values of the growth equation and other life parameters calculated in this study are important information for the management of the artisanal fishery in the Central Pacific coast of Mexico. Results have been compared with other species of the same family and M. brevipinnis of years ago, which validates and gives certainty to its use in future models.
ACKNOWLEDGMENTS
Artisanal fishers from Jalisco were very helpful with their information and facilitating us organisms for samples; also MS Alejandro Gómez-Ponce and Biol. Claudio Padilla-Hernández for their help in the lab. Several institutions gave financial support: Universidad Nacional Autónoma de México, CONACyT and INAPESCA.
EEB, MGC, KGF, EGCS, AGB and MPG participated in the collection, sampling and analysis of the fish. We all also participated in the data analysis and growth parameters calculations. The MGC and EEB wrote the paper.
REFERENCES
- Aguirre-Leon, A. and S. Diaz-Ruiz, 2006. Estructura de tallas, madurez gonadica y alimentacion del pez Diapterus rhombeus (Gerreidae) en el sistema fluvio-deltaico Pom-Atasta, Campeche, Mexico. Rev. Biol. Trop., 54: 599-611.
Direct Link - Appeldoorn, R.S., 1992. Interspecific relationships between growth parameters, with application to haemulid fishes. Proceedings 7th International Coral Reef Symposium, June 22-27, Guam, Micronesia, pp: 899-904.
Direct Link - Espino-Barr, E., M. Gallardo-Cabello, F. Gonzalez-Orozco and A. Garcia-Boa, 2004. Analisis del crecimiento y la mortalidad de Anisotremus interruptus (Gill, 1863) (Perciformes: Haemulidae) en la costa de Colima, Mexico. Bol. Invest. Mar. Cost., 33: 67-75.
Direct Link - Espino-Barr, E., M. Gallardo-Cabello, A. Garcia-Boa, E.G. Cabral-Solis and M. Puente-Gomez, 2006. Morphologic and morphometric analysis and growth rings identification of otoliths: Sagitta, asteriscus and lapillus of Caranx caninus (Pisces: Carangidae) in the coast of Colima, Mexico. J. Fish. Aquat. Sci., 1: 157-170.
CrossRefDirect Link - Espino-Barr, E., M.G. Cabello, E.G. Solis, A.G. Boa and M.P. Gomez, 2008. Growth of the Pacific jack Caranx caninus (Pisces: Carangidae) from the coast of Colima, Mexico. Rev. Biol. Trop., 56: 171-179.
PubMedDirect Link - Ford, E., 1933. An account of the herring investigations conducted at Plymouth during the years from 1924 to 1933. J. Mar. Biol. Assoc. UK., 19: 305-384.
CrossRefDirect Link - Granados-Flores, K., M. Gallardo-Cabello, E. Espino-Barr and E.G. Cabral-Solis, 2010. Age determination of Microlepidotus brevipinnis (Steindachner, 1869) (Pisces: Haemulidae) in the coast of Jalisco, Mexico, by reading otoliths and scales. Int. J. Zool. Res., 6: 1-12.
CrossRefDirect Link - Lucano-Ramirez, G., S. Ruiz-Ramirez and J.A. Rojo-Vazquez, 2006. Composicion por tallas y ciclo reproductivo de Pseudupeneus grandisquamis (Pisces: Mullidae) en el Pacifico Central Mexicano. Revista de Biologia Trop., 54: 195-207.
Direct Link - Munro, J.L. and D. McB. Williams, 1985. Assessment and management of coral reef fisheries: Biological, environmental and socio-economic aspects. Proc. Int. Coral Reef Congr. Tahiti, 4: 545-578.
Direct Link - Nelson, J.S., E.J. Crossman, H. Espinosa-Perez, L.T. Findley, C.R. Gilbert, R.N. Lea and J.D. Williams, 2004. Common and Scientific Names of Fishes from the United States, Canada and Mexico. 6th Edn., American Fisheries Society, USA., ISBN-10:1888569670, pp: 386.
Direct Link - Pajuelo, J.G., J.M. Lorenzo and M. Gregoire, 2003. Age and growth of the bastard grunt (Pomadasys incisus: Haemulidae) inhabiting the Canarian archipelago, Northwest Africa. Fish. Bull., 101: 851-859.
Direct Link - Pauly, D., 1978. A preliminary compilation of fish length growth parameters. Berlin Inst. Meereskunde Christian-Albrechts-Univ. Kiel, 55: 1-200.
Direct Link - Rojas, J.R., M.E. Maravilla and F. Chicas, 2004. Habitos alimentarios del pargo mancha Lutjanus guttatus (Pisces: Lutjanidae) en Los Cobanos y Puerto La Libertad, El Salvador. Rev. Biol. Trop., 52: 163-170.
Direct Link - Rojas-Herrera, A.A., M. Mascaro and X. Chiappa-Carrara, 2004. Habitos alimentarios de los peces Lutjanus peru y Lutjanus guttatus (Pisces: Lutjanidae) en Guerrero, Mexico. Rev. Biol. Trop., 52: 959-972.
Direct Link - Safran, P., 1992. Theoretical analysis of the weight-length relationship in fish juveniles. Mar. Biol., 112: 545-551.
CrossRef - Saucedo-Lozano, M., G. Gonzalez-Sanson and X. Chiappa-Carrara, 1999. Alimentacion natural de juveniles de Lutjanus peru (Nichols y Murphy, 1922) (Lutjanidae: Perciformes) en la costa de Jalisco y Colima, Mexico. Ciencias Marinas, 25: 381-400.
Direct Link - Sauskan, V.I. and A. Olayechea, 1974. Biology and population abundance of the tomtate (Haemulon aurolineatum Cuvier and Valenciennes, 1830, Pomadasyidae, Perciformes) on Campeche Bank. Soviet-Cuban Fish. Invest., 4: 135-150.
Direct Link - Taylor, C.C., 1960. Temperature, growth and mortality-the Pacific cockle. J. Conseil., 26: 117-124.
CrossRef - Von Bertalanffy, L., 1938. A quantitative theory of organic growth (Inquiries on growth laws. II). Hum. Biol., 10: 181-213.
Direct Link - Walford, L.A., 1946. A new graphic method of describing the growth of animals. Biol. Bull., 90: 141-147.
PubMedDirect Link - Allen, G.R. and D.R. Robertson, 1994. Fishes of the Tropical Eastern Pacific. 1st Edn., University of Hawaii Press, USA., ISBN-10: 0824816757.
Direct Link