
M. Yousefian
Ecological Institute of Caspian Sea, P.O. Box 961, Sari, Iran
A. Nejati
Ecological Institute of Caspian Sea, P.O. Box 961, Sari, Iran
Journal of Fisheries and Aquatic Science
Year: 2008 | Volume: 3 | Issue: 6 | Page No.: 384-391







ABSTRACT
In this study, inbreeding depression and abnormality effects on the basis of differences between inbred and outbred half-sib family groups of rainbow trout, Oncorhynchus mykiss are investigated. One generation of brother-sister mating, inbreeding depression in rainbow trout was calculated for three production traits, percentage egg hatchability, percentage survival of fry at active nutrition stage and weights at 150, 225 and 300 days. The inbreeding depression obtained for half-sib mating was 1.25 for percentage hatchability, 2.9 for percentage fry survival and up to 28.1, 35.2 and 35.6 for percent weight at 150, 225 and 300 days, respectively. Egg hatchability, fry survival and weight at early growth stage were not significant while a highly significant inbreeding depression was found for weight at 300 day (p<0). The difference between inbreeding depression for male and female half-sib group was non significant (p≥0.05). Although the inbreeding embryo had a noticeable though not significant negative effect on survival larval stage, it had a strong and significantly negative effect of growth rate at later stages of development. In the second series of experiments, the three traits showed the same pattern as the first one. Following one generation of full-sib mating, the calculated effect of inbreeding on the number of live fish and total weight of fish at 150 days of age showed a reduction of 5.83 to 8.49 weight percent in 2 groups. About 23% of larvae at hatching showed abnormal shapes of bighead, curved body, shortened tail and big abdomen, etc. There was frequently a type of axial deformations that were related to notochord alternations during embryogenesis. About 19% of abnormal larvae survived only upto feeding stage but 4% survived and reached juvenile and adults stage.
PDF Abstract XML References Citation
How to cite this article
M. Yousefian and A. Nejati, 2008. Inbreeding Depression by Family Matching in Rainbow Trout (Oncorhynchus mykiss). Journal of Fisheries and Aquatic Science, 3: 384-391.
DOI: 10.3923/jfas.2008.384.391
URL: https://scialert.net/abstract/?doi=jfas.2008.384.391
DOI: 10.3923/jfas.2008.384.391
URL: https://scialert.net/abstract/?doi=jfas.2008.384.391
INTRODUCTION
Rainbow trout is a species of general importance in Iranian Aquaculture. Iran has many rivers and springs which provide sources of cold water, favorable for the growth of rainbow trout (Oncorhynchus mykiss). Using these natural advantages, commercial trout farming increased in recent years and the production was about 34000 ton in 2005. Production of this species expanding very fast and most of the farmers try to use the fry from their own brood stocks. Rainbow trout have advantage for use in breeding, notably: with the small number of breeding individuals all the progeny of succeeding generations can be produced but this leads to increased inbreeding. Moreover, the practice of mating unequal number of males and females, usually in this case, a smaller number of males than females, also increased the probability of inbreeding (Falconer, 1989). Inbreeding is mating between individuals which are related to each other by ancestry. Inbreeding depression is the effect of inbreeding measured as the decline in performance of a quantitative trail. It has been investigated in most domestic and laboratory animal species including fish.
Studies to measure depression at a series of inbreeding levels in rainbow trout populations using the method of inbred-outbred half-sib families are discussed. While the actual depression estimates varied widely between populations and inbreeding levels, significant levels of depression were found in many hatchery, field and brood stock performance traits after only one generation of brother-sister mating (Kincaid, 1983).
Therefore, the high level of abnormal fish especially spinal malformation appearing in hatchery fish that is an important problem in many private farms. Malformation often is associated with growth depression, leading to high mortality rates at early fry stage. Gjerde et al. (1983) investigated the effect of three levels of inbreeding (F = 0.25, 0.375 and 0.50) on survival and growth, found that inbreeding caused significant depression in survival of eyed eggs, alevins and fry, in growth rate of adults but not in growth rate upto 160 days after first feeding. The source and the rate of abnormality still is a problem for many hatchery farms.
The objective of this study was to investigate the magnitude effect of the inbreeding on survival rate, body weight and abnormalities in brother-sister mating of rainbow trout in one of the hatcheries in Iran.
MATERIALS AND METHODS
The rainbow trout breeders were obtained from a captive brood stock, fed special (Ecogen 13, Biomar, Fr.) pellets contained, crude protein 50%, crude fat 13%, ash 9.5%, fiber 1.9% and P total 1.3%. The breeders maintained by Kelardasht salmon propagation center (Iran), one pair from Yasoj fish farm (Center part of Iran-group A) and one pair from Kelardasht salmon propagation center (North part of Iran-group B). The breeders were in the same age group (3 years) and were selected based on similar growth rate, 2.8 kg. The experiment conducted from 2001-2005.
A random block design was done between A and B groups using 4 randomly chosen sires and 4 randomly chosen dams from each groups. In second experiment a full factorial diallele cross was done between A and B groups using 4 randomly chosen sires and 4 randomly chosen dams from each group, thus theoretically producing (4+)(4+4) = 64 half-sibs and full-sibs families. All dam and sire age were equal, thus making the dam and sire effect possible. Sixteen full-sib and half-sib mating in experiment one and the 64 mating in experiment 2 were individually incubated in a fully randomized design of eighty standard California incubators. Incubation went on at 8±0.5°C until hatching around 420° days (°D) and then until swim-up at 750° days (°D). After hatching the fish were transferred to nursery tanks, one tank for each full-sib family of 150 fry (1 g L-1). The tanks were rectangular of 2x0.5x0.2 m, receiving about 12 L min-1 of fully oxygenated water. Feeding was initiated when the fry showed signs of swimming. Fifty days of age, the fish were transferred to a set of 1.5x1.5x1 m concrete pond, with a volume of 2000 L receiving about 30 L min-1 of fully oxygenated water. Feeding was provided by hand feeding at a rate of 1.5% b.wt. day-1. Water temperature ranged from 8-12°C with an average 11±1.41°C.
Data Collection
The following measurements were taken.
| • | Fertilization rate |
| • | Survival from fertilization to eyed stage and from eyed stage to swim-up |
| • | Hatchability rate |
| • | Percentage of swim up fry |
| • | Percentage of active feeding stage |
| • | Growth at different period of time |
For estimate the fertilization and eyed eggs rate 100 eggs were tested. Data of survival rate were estimate by the daily decrease in the number of larvae during experiment period (300 days).
Abnormal Larvae and Juvenile
The percentage of deformed specimens was estimated by counting 100 larvae for each experimental period. In progressive stages normal and lordotic (n = 60) juveniles were anaesthetized, weighted and radiographed using a medical x-ray system. The system was adjusted for fish at the size of 30-60 g. The x-ray pictures were used for examination of the skeleton.
The lordosis was evaluated by measuring the place and number of vertebrate at which defect occur.
Statistical Analysis
Statistical comparisons between survival data were performed with one-way factorial analysis (ANOVA), with confidence level of 95%, followed by the fisher, Protected Least Significant Difference, (PLSD) multiple comparison test using the program SPSS 11.5 for Windows.
The weight gain was measured for 10 fish in each treatment. The percentage of deformed larvae was estimated by counting total larvae for each experimental period.
RESULTS
The mean number of fertilized ova, eyed eggs, survival at hatching stage and swim up is shown in Table 1.
The analysis of variance showed that the effect of inbreeding was not significant on fertilization, eyed stage hatching stage, neither at the swim-up nor at active feeding stage (p>0.05).
Analysis of variance revealed that effect of inbreeding on the weight of juvenile was not significant at 150 days and 225 days weight giant of fishes but a highly significant effect was at 300 days of rearing (p<0.00, Table 1). The difference between inbreeding depression for male and female half-sib group was non significant (p>0.05). The differences were not significant of sire line (p<0.111) and dam line (p<0.819) too.
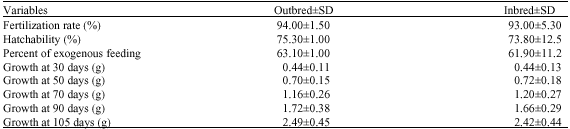
In second series of experiment sampling at fertilization, active feeding stage and weight gain at 30, 50, 70, 90 and 105 were done. Result revealed (Table 2), as in the first experiment that effect of inbreeding at any developmental stages of growing from 30 to 105 days (p<0.39) and all other effects such as sire line or dam line were non significant.
| Table 1: | Comparison of the half-sib (ANOVA) on rainbow trout survival and 150-300 days growing |
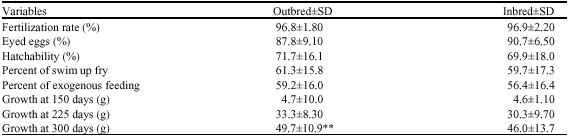 | |
| **represent significant difference at (p<0.01) | |
| Table 2: | Comparison of the half-sib (ANOVA) on rainbow trout survival and 30-105 days growing |
 | |
| Table 3: | Calculated inbreeding depression in weight of fish and number of fish at 159 days of age due to the combined joint effects of egg hatchability, fry survival (exogenous feeding stage) and attained weights at F = 0.25 |
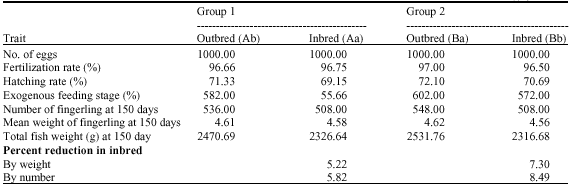 | |
A and B refer to Yasoj and Kelardasht fish farm females and a and b to males of two stocks | |
The accumulative effect of inbreeding depression of fry survival and 150 days weights on resulting total weight and total number of fish produced at 150 days of age, was calculated by using the mean inbreeding effects measured at the F = 0.25 level of inbreeding in Table 3.
The combined effects of the remaining two traits at an inbreeding level of full-sib mating reduced the total weight of fish produced at 150 days of age 5.83 and 8.49% in two groups.
Abnormal Larvae and Juvenile
Result on the effect of inbreeding on growth related variable at active feeding and weight at 150-300 days showed non-significant difference between percentage of abnormal larvae and juvenile between half sibs and full fish mating (p≥0.05).
The highest mortality occurred during one month at the time of hatching up to feeding stage. Many abnormal larvae could not rupture the eggs shell and died before hatching (18.4%). A few hatched but was not able to swim; they stayed at bottom of incubator (10.3%). Others reabsorbed the yolk sac and start internal feeding.
On the 30th day, the percentage of larvae survival was 57.8%; this figure decreased to 54.9%, five month later, when larvae weighted 4.6 g. The most frequent observed deformations at this time were deformation of vertebral column in tail (up to 92% of abnormal fry) and other abnormalities observed were operculum atrophies (5%), cranial abnormalities (1%) and abnormal shape.
About 10% of the newly hatched larvae observed, displayed clearly distinguishable skeletal deformities. Six days after hatching, yolk sac reabsorption occurred; larvae become active and commenced feeding. The number of deformed larvae progressing decreased from this period and at the end of larval period, they represented about 3% of survival larvae. Most of these deformed animals had deformation in the vertebral column.
Abnormal specimens showed unbalanced growth, which started to be clearly noticeable 3-4 week after hatching (1 g). The difference progressively increased and, 150 days after hatching, the juveniles measured 14.1 cm in length, while normal specimens measured 12.4 cm. X-ray examinations shows the place of defect (Fig. 1) and some juvenile showed V-shaped vertebral column. In some juveniles a skeletal sign of deformity more than one point of curvature was identified, whereas others showed such slight undulating curvatures of the vertebral column, that externally they were recognized only by the irregular trajectory of this lateral line. Juvenile salmo trout have 63 spinal vertebral (21 cervical, 21 abdominal and 20-22 caudal). The frequency of affected spinal vertebrae was estimated and the vertebrae between 38-48 were found to be most frequently affected with a maximum frequency at the 43rd vertebra. The lowest frequency was found to correspond to the vertebrae behind the head.
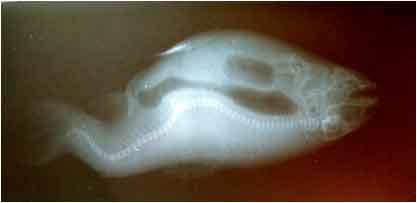 | |
| Fig. 1: | Deformation in vertebral column of rainbow trout is shown by radiograph assay |
DISCUSSION
There have been reports of high rate of occurrence of abnormal juveniles and adults (up to 20%) from several trout farms in Iran.
In the present investigation, environmental conditions seemed to have caused higher rate of abnormalities than inbreeding as the period of incubation was very long and the eggs of cold water fish are more sensitive to environmental variables.
In reared rainbow trout, inbreeding was thought to involve two conditions, abnormality and reduced growth rate. The accumulation of inbreeding is a natural consequence for closed population undergoing artificial selection.
Critical values for inbreeding level have not been established in fish, but Tave (1993) suggested using F = 5% as a conservative estimate and F = 10% as a tolerate estimate. In the present study, detectable harmful effects on important traits such as fecundity, survival and growth at an inbreeding level of 0.25 have been investigated. For this purpose the effect of inbreeding was estimated in male and female half-sib families.
This experiment demonstrated reduced survival at eyed stage, swim-up and at feeding stage in male and female half-sib, but difference in survival rate observed was not significant as was reported by (Kincaid, 1983) for Rainbow trout. During the rearing period from 150 to 300 days (rearing at 10°C) and from 30 to 105 days (at 18°C) at advanced stage of rearing weight reduction due to inbreeding was significant.
This trend was expected considering that these measurements were taken at a series of age intervals on the same families. Since growth occurs as a function of body mass at the beginning of the particular growth period, any deviation between the inbred and outbred, weight at the end of one growth period would be expected to increase, in each subsequent growth period (Kincaid, 1983). Therefore at progressive growth the mean depression in weight showed significant difference among male and female half-sib with full-sib mating. The cumulative effect of inbreeding depression of fry survival and weight at 150 days, showed an average 7.16% reduction in full-sib groups. This reduction evident at the fingerling stage (4.5 g) was found to be more pronounced and significant at the juvenile stage. Inbreeding depression in body weight showed a clear trend to increasing trend with age from 150 days to adult stage. It may be due to the cumulative effect of inbreeding as the body of the fish successively grows larger as stated by Su et al. (1996a).
Inbreeding depression could not be found in body weight at early stages before 256 days age, as stated by Su et al. (1996a). They stated that at relative low level of inbreeding (an average up to 19.5%) inbreeding depression in body weight at early age stages is not serious enough to be detectable. It is in accordance with the result obtained at inbreeding level of F = 0.25. Gjerde et al. (1983) found no significant inbreeding depression in growth of fingerlings even at high level of inbreeding (F = 0.25-0.50) and they assumed that the observed inbreeding depression of survival counteracted inbreeding depression in body weight because mortality may preferentially act on the smallest individuals in the population.
The effects of inbreeding of the maternal and paternal family groups could be markedly different or even significant. Maternal effect was expected to be appreciably larger than paternal effect during early period of life because of the influence of embryo size and the quantity and quality of yolk materials, which are almost entirely determined by the female during egg formation. The formation of maternal half-sib families from a random half of the eggs of a single female maximized the uniformity of the eggs used in each of the female mating. In parental families, however, the range in size and general quality of the eggs varied at random, depending on the particular females selected. Inbreeding would be expected to have a depression effect on maternal influence where the female was herself inbred (Falconer, 1989). Using large population of breeders (more than 250 pairs) in Kelardasht fish farm and choosing randomly few breaders, no inbred male or female would be expected in half – sib mating or control groups.
In other hand as Su et al. (1996b) reported, maternal effects should not be large for salmonid because the offspring are not nurtured. The most likely source of maternal effect would be difference in egg size and egg quality. However as reported by Crandell and Gall (1993), in most studies with rainbow trout, any correlation observed between egg size and early growth disappears soon after first feeding. Therefore full-sib family effects, as observed in the present study, most likely reflect only common environmental effects. The same statement was obtained by Su et al. (1996b) in their study in rainbow trout. Existing studies shows dam effect on survival at eyed stage, whereas swim–up and active feeding stage reveal no significant effect. Egg size may show significant effect in salmonid as observed in many studies (Rana, 1985; Marteinsdottir and Albe, 1992; Wallace and Aasjord, 1984). However as shown by Vanderputte et al. (2002) and Jonsson and Sravarsson (2000), egg size had no effect on the survival from fertilization to the eyed stage. A review of studies of the effect of sire or dam revealed that survival is related to environmental factors or at least can be considered to have a low additive genetic component (Kanis et al., 1976; Kinghorn, 1983).
In a review of several reports, Gjedrem (1992) found that heritabilities estimated from dam components were a bit higher than those estimated from sire components of variance, suggesting strong common environmental and/or non-additive genetic effect.
The full-sib family effects on body weight at two experiments (30-105 and 150-300) showed two differences in trends. First, the percentage inbreeding depression did not increase as age increased from 30 to 105 days. This trend is an anomaly largely explained by the higher fry mortality observed in inbred lots, which lowered fish density and thus reduced competition for the available space and food, even though thinning was done randomly, by culling extremely small fish, resulting in a decrease in the variance within full-sib families. The growth of the remaining fry reared at a lower fish density would give the appearance of decreasing effect of inbreeding depression, as suggested by the trend seen in this experiment and those of Su et al. (1996a). Second, markedly evident of increasing depression with advancing age was seen from 150 to 300 days. A relatively large and significant increase of this effect of depression was found at 300 days.
Inbreeding depression for body weight was relatively high at 300 days of age. Inbreeding effect as well as inbreeding rate was high for body weight at harvest time (Pante et al., 2001a). This may be due to some families responding more severely to inbreeding or selected future parents might have come from related families (Pante et al., 2001b).
As we stated earlier, the high level of spinal malformations appearing in hatchery rainbow trout is an important problem in fish farms. Abnormal fish have an unfavorable shape, decreasing the production and increasing the cost.
The etiology of these syndromes in fish has been study by many researchers and nutritional, pathogenic, environmental and genetic causes have been cited. Nutritional factors such as deficiencies in ascorbic acid, vitamins and triptophem in the diet of parental fish and larvae have shown different skeletal deformities (Walton et al., 1984). Pathogens such as the mynosporidian Myxosoma sp. Andrades et al. (1992) may induce congenital or postnatal skeletal deformation in fish. Environmental factors such as density of eggs, mechanical or thermal shocks, pollution, radiation, salinity, oxygen deficiency of water and light intensity have also been reported to cause aberrations in development (Madsen and Dalsgaard, 1999; Sloof, 1982; Wiegand et al., 1989).
In the present study no significant difference in the number of abnormal fish at active feeding stage at the end of yolk resumption or in the growth at 150, 225 and 300 days was observed. Any markedly significant difference between different mating to confirm genetic causes of variability were also not observed during the investigation. In spite of high percent (about 10%) of larvae showed abnormalities and the rate decreased to 3% on the 300th day with most of the abnormal juveniles exhibiting vertebral abnormalities in the tail region. Survival of the larvae with vertebral abnormalities was found to be higher compared to other more serious types of abnormalities.
Therefore, in contrast to the other abnormalities, vertebral abnormality in larvae does not totally interfere with feeding, but during culture they are collected and culled at different ages. In these respect economical losses of vertebral abnormality is higher than those which lead to larval mortality at the time of hatching. To avoid losses and to achieve optimum production and income it is suggested that rainbow trout culture should be carried out only by standard farms using good quality of water, high value nutrition, large number of breeders and avoiding of full or half-sib mating through identification of individual families.
REFERENCES
- Andrades, J.A., J. Becerra and P. Fernandez-Llebrez, 1996. Skeletal deformities in larval, juvenile and adult stages of cultured gilthead sea bream (Sparus aurata L.). Aquaculture, 141: 1-11.
CrossRef - Crandell, P.A. and G.A.E. Gall, 1993. The genetics of body weight and on early maturity based on individually tagged rainbow trout (Oncorhynchus mykiss). Aquaculture, 117: 77-93.
CrossRef - Falconer, D.S., 1989. Introduction to Quantitative Genetics. 3rd Edn., Longman Scientific and Technical, New York, USA Pages: 438.
Direct Link - Gjerde, B., K. Gunnes and T. Gjedrem, 1983. Effect of inbreeding on survival and growth in rainbow trout. Aquaculture, 34: 327-332.
CrossRef - Jonsson, B. and E. Svavarsson, 2000. Connection between egg size and early mortality in arctic charr, Saluelinus alpinus. Aquaculture, 187: 315-317.
CrossRef - Kanis, E., T. Refstie and T. Gjedrem, 1976. A genetic analysis of egg, alevin and fry mortality in salmon, sea trout and rainbow trout. Aquaculture, 8: 259-268.
CrossRef - Kincaid, H.L., 1983. Inbreeding in fish populations used for aquaculture. Aquaculture, 33: 215-227.
CrossRef - Kinghorn, B.P., 1983. A review of quantitative genetics in fish breeding. Aquaculture, 31: 283-304.
CrossRef - Marteinsdottir, G. and K.W. Able, 1992. Influence off egg size on embryos and larvae of Fundulus heetroclitus (L.). J. Fish Biol., 41: 883-896.
CrossRef - Pante, M.J.R., B. Gjerde and I. McMillan, 2001. Effect of inbreeding on body weight at harvest in rainbow trout. Oncorhynchus mykiss. Aquaculture, 192: 201-211.
CrossRef - Pante, M.J.R., B. Gjerde and I. McMillan, 2001. Inbreeding levels in selected populations of rainbow trout, Oncorhynchus mykiss. Aquaculture, 192: 213-224.
CrossRefDirect Link - Rana, K.J., 1985. Influence off egg size on growth, onset of feeding, point-of-no-return, and survival of unfed Oreochromis mossambicus fry. Aquaculture, 46: 119-131.
CrossRef - Madsen, L. and I. Dalsgaard, 1999. Vertebral column deformities in farmed rainbow trout (Oncorhynchus mykiss). Aquacult., 121: 41-48.
CrossRef - Sloof, W., 1982. Skeletal anomalies in fish from polluted surface waters. Aquat. Toxicol., 2: 157-173.
CrossRef - Su, G.S., L.E. Liljedahl and G.A.E. Gall, 1996. Genetic and environmental variation of body weight in rainbow trout (Oncorhynchus mykiss). Aquaculture, 144: 71-80.
CrossRef - Su, G.S., L.E. Liljedahl and G.A.E. Gall, 1996. Effects of inbreeding on growth and reproductive traits in rainbow trout (Oncorhynchus mykiss). Aquaculture, 142: 139-148.
CrossRef - Vanderputte, M., E. Quillet and B. Chevassus, 2002. Early development and survival in brown trout (Salmo trutta fario L.): Indirect effects of selection for growth rate and estimation of genetic parameters. Aquaculture, 204: 435-445.
CrossRef - Wallace, J.C. and D. Aasjord, 1984. An investigation of the consequence of egg size for the culture of Arctic charr, Saluelinus alpinus (L.). J. Fish Biol., 24: 427-435.
CrossRef - Walton, M.J., R.M. Coloso, C.B. Cowey, J.W. Andron and D. Knox, 1984. The effects of dietary tryptophan levels on growth and metabolism of rainbow trout (Salmo gairdneri). Br. J. Nutr., 51: 279-288.
CrossRefDirect Link - Wiegand, M.D., J.M. Hataley, C.L. Kitchen and L.C. Buchanan, 1989. Lnduction of developmental abnormalities in larval goldfish, Carassius auratus L., under cool incubations. J. Fish Biol., 35: 85-95.
CrossRef