
Elaine Espino-Barr
CRIP, Manzanillo, INP. Playa Ventanas, Colima, Mexico
Manuel Gallardo-Cabello
UNAM, Mexico, D.F
Arturo Garcia-Boa
CRIP, Manzanillo, INP. Playa Ventanas, Colima, Mexico
Esther G. Cabral-Solis
CRIP, Manzanillo, INP. Playa Ventanas, Colima, Mexico
Marcos Puente-Gomez
CRIP, Manzanillo, INP. Playa Ventanas, Colima, Mexico
Journal of Fisheries and Aquatic Science
Year: 2006 | Volume: 1 | Issue: 2 | Page No.: 157-170







ABSTRACT
This study describes and provides a morphometric analysis of the otoliths sagittae, asterisci and lapilli of the Pacific jack Caranx caninus Günther, 1869, in the tropical Mexican Pacific. The relationship among length, width and weight of the sagittae is expressed by the following equations: y = 0.772x0.950 (rostrum-antirostrum), y = 0.480x0.791 (rostrum-width) and y = 0.00006x2.595 (rostrum-weight). In the case of the asterisci: y = 1.704x1.077 (length-width) and y = 0.00048x2.478 (length-weight). For the lapilli y = 1.047x0.949 (length-width) and y = 0.00028x3.047 (length-weight). The growth of these otoliths was also related to the length of the fish. The average length was calculated for each of the six growth rings identified in the sagittae and the asterisci; the results are: 0 = 6.33 cm, 1 = 15.09 cm, 2 = 32.19 cm, 3 = 41.50 cm, 4 = 48.17 cm, 5 = 55.58 cm and 6 = 58.75 cm.
PDF Abstract XML References
How to cite this article
Elaine Espino-Barr, Manuel Gallardo-Cabello, Arturo Garcia-Boa, Esther G. Cabral-Solis and Marcos Puente-Gomez, 2006. Morphologic and Morphometric Analysis and Growth Rings Identification of Otoliths: Sagitta, Asteriscus and Lapillus of Caranx caninus (Pisces: Carangidae) in the Coast of Colima, Mexico. Journal of Fisheries and Aquatic Science, 1: 157-170.
DOI: 10.3923/jfas.2006.157.170
URL: https://scialert.net/abstract/?doi=jfas.2006.157.170
DOI: 10.3923/jfas.2006.157.170
URL: https://scialert.net/abstract/?doi=jfas.2006.157.170
INTRODUCTION
The identification of the growth rings is commonly made through the analysis of the sagittae because it is the largest otolith and therefore easier to extract and to examine (Campana and Neilson, 1985; Stevensen and Campana, 1992; Mascareña-Osorio et al., 2003; Francis et al., 2005; Begg et al., 2005; Berg et al., 2005).
Some authors (Victor and Brothers, 1982; Brothers et al., 1983; Solomon et al., 1985; Suthers et al., 1989; David et al., 1994) have determined the daily growth increments in the lapilli of larvae and juvenile specimens. Barkman (1978), Bolz and Lough (1983 and 1988), Lagardere (1989) and David et al. (1994) have used the asterisci with the same purpose.
However, there are no studies on age determination on periodical growth for adult fish analyzing the rings of their asterisci and lapilli. For the same reason, the morphology and the morphometry of these otoliths has not been studied.
This study is a morphologic and morphometric analysis of the otoliths: sagittae, asterisci and lapilli of the Pacific jack Caranx caninus, as well as the identification of the growth rings in the sagittae and the asterisci.
The Pacific jack Caranx caninus Günther, 1867, is a coastal pelagic species of commercial value in the artisanal fishery of the tropical Mexican Pacific. These species are distributed from the coasts of Mexico to the North of Perú (Chirichigno et al., 1982; Fisher et al., 1995). An average of 76 tons are fished annually in the state of Colima which is used for human consumption. There are studies on their taxonomic description (Castro-Aguirre et al., 1999; Chirichigno et al., 1982; Fisher et al., 1995) and on biological and fishery information (Cruz-Romero et al., 1993; Espino-Barr, 2000; Espino-Barr et al., 2003, 2004). Torres-Aguilar (2002) made the identification of the growth rings of this species through the analysis of spines. However, there are no data on the analysis of the growth rings in otoliths. The scales of this species are replaced during its growth, so the otoliths are one of the adequate structures for the age determination analysis.
MATERIALS AND METHODS
Fifty specimens of C. caninus were obtained monthly between January and December 2002 from the commercial catches of the artisanal fishery of Manzanillo, Colima (Mexican Pacific). In order to obtain every length and age groups, sampling method included the individuals captured with pound net, beach seine, hand line and gillnets. The variables registered for each organism were Standard Length (SL), weight and sex.
The otoliths sagittae, asterisci and lapilli were obtained through a traverse section in the ventral cranial cavity, the brain was removed and the right and left semicircular canals were extracted. The otoliths were rinsed with water and stored dry in Eppendorf tubes labeled with number, date, standard length and sex.
Scanning photographs of the structure and microstructure of the otoliths were obtained with a scanning electronic microscope, from the Institute of Physics of the Universidad Nacional Autónoma de México.
The terminology of the glossary of Secor et al. (1992) was employed for the description of the labyrinth system and the sagittae. Similar concepts were applied to the description of the asterisci and lapilli which have not been reported before.
The otolith sample size was calculated with the formula described by Daniels (1991) and through the observation in a stereoscopic microscope with a graduated ocular lens; data on the length and the width for each otolith were registered (Fig. 4, 9 and 10). The weight of each otolith was also registered in an ultranalytic scale with a precision of a ten thousandth gram. In order to eliminate possible differences due to variations between the otoliths in the same individual, all measures were obtained from the right-sided sagittae, asterisci and lapilli.
Constants of the relationships of the sagittae were calculated for Rostrum Length (RL), Antirostrum Length (AL), Width (Wi) and Weight (We). For the asterisci and lapilli the indexes of the constants of the relationship were obtained for Length (L), Width (Wi) and Weight (We). The relationships between the standard length of the fish and all the measures of the three otoliths were also made. The regressions were done by the least squares. For the evaluation of the relations and the analysis of the possible morphometric differences between the otoliths of males and females, a variance analysis (anova) was carried out (Mendenhall, 1987; Zar, 1996).
The growth rings were identified, observing the sagittae and the asterisci in the stereoscopic microscope with transmitted light and the average size of each ring was calculated.
RESULTS AND DISCUSSION
Labyrinth System of C. caninus
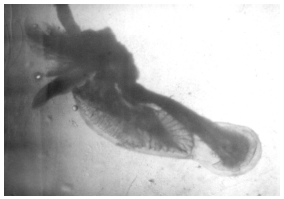
The membranous labyrinth of C. caninus is formed by a sac-like tissue where the semicircular canals end in a main chamber: the vestibular apparatus which is formed by dorsal sacs, the pars superior (where lapilli are contained) and ventral sacs, the pars inferior (where sagittae and asterisci are enclosed). The individual vestibules which enclose these three pairs of otoliths are the utriculus, the sacculus and the lagena which contain the lapillus (Fig. 1), the sagitta and the asteriscus (Fig. 2), respectively (Lagler et al., 1962; Secor et al., 1992).
The pars superior is responsible for the equilibrium and the pars inferior is related with the perception of sound, angular and gravity acceleration (Lowenstein, 1957; Holst et al., 1950).
Sensorial nervous cells (neuromast cells) are present in each cavity giving support to the otoliths; there are in contact with the macula through which the growth nutrients are deposited and they transmit the stimuli to the brain through the eighth cranial nerve (Fig. 3) (Mugiya, 1964, 1966 a,b).
Otoliths (sagittae) are structures formed by otoline, a high weight molecular protein (Degens et al., 1969) and calcic carbonate (Lagler et al., 1962) in the form of aragonite (Hickling, 1931; Brandenberger and Schintz, 1945; Sasaki and Miyata, 1955; Carlström, 1963; Gallardo-Cabello, 1986b).
The sagitta is the largest otolith in Caranx caninus, reaching a total length of 7.41 mm; the lapillus measures 2.98 mm and the asteriscus 1.37 mm, in organisms of 30 cm of standard length.
Description of the Otoliths of the Pacific Jack Caranx caninus
Description of the Sagitta
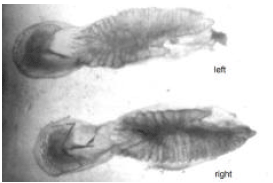
The anterior margin exhibits a sharp “excisura major” and “antirostrum” which tend to disappear as the organism grows showing a small protuberance in the medium dorsal margin of the older fishes (Fig. 4). The “rostrum” shows in some cases, small perforations in one of its otoliths and morphologic variations, that make the right sagitta different from the left one.
The posterior margin shows a rounded “postrostrum” without “excisura minor” or “pararostrum” (Fig. 5).
The anterior, posterior, ventral and part of the dorsal margin show regular denticles in the form of a saw and the dorsal margin which corresponds to the rectilinear antirostrum (Fig. 4).
The internal aspect of the otolith is convex, a characteristic which increases as fish ages; a deep sulcus runs over its smooth surface which is not differentiated between the ostium and the cauda (Fig. 6).
The base of the acoustic channel increases in width from the center to the end of the otolith, it is made of calcic carbonate crystals oriented in an epitaxial growth and showing diverse orientation angles (Fig. 7).
The external aspect is concave; its thickness decreases longitudinally from the middle part of the otolith to its anterior margin.
Figure 8 shows the growth bands formed from the otolith core, these rings start to appear as soon as the larvae hatch and the vitelin sack (egg yolk) has been absorbed (Brothers et al., 1976; Struksaker and Uchiyama, 1976). The bands which are constituted from the core to the otolith margins represent different growth rate patterns that may occur daily and seasonally and makes it possible to identify the growth rings through which age groups can be obtained (Hickling, 1931; Dannevig, 1956; Kelly and Barker, 1961; Degens et al., 1969; Pannella, 1971; Gallardo-Cabello, 1986 a,b).
Description of the Asteriscus
The anterior margin of this otolith is a sharp triangular structure that divides the asteriscus, which is a fan-like structure with sides: the dorsal side and a smaller ventral side.
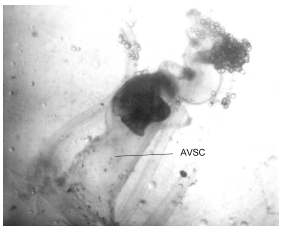 | |
| Fig. 1: | Lapillus contained in the utriculus and section of the anterior vertical semicircular canal(AVSC) of the membranous labyrinth in an individual of Caranx caninus (35 cm of standard length) (increased 15 times) |
 | |
| Fig. 2: | Sagittae contained in the sacculus and the asterisci in the lagena of the membranous labyrinth in an individual of Caranx caninus (35 cm standard length) (increased 10 times) |
 | |
| Fig. 3: | Section of the right membranous labyrinth showing the macula on the sagitta and asteriscus in an individual of Caranx caninus (35 cm of standard longitude) (increased 10 times) |
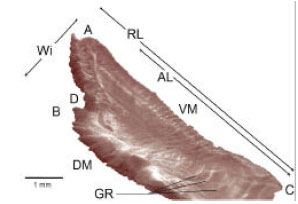 | |
| Fig. 4: | Main characteristics of the right sagitta, external aspect of Caranx caninus: A = rostrum, B = antirostrum, C = postrostrum, D = excisura major, DM = dorsal margin, VM = ventral margin, AL = antirostrum length, RL = rostrum length, Wi = width, GR = growth rings |
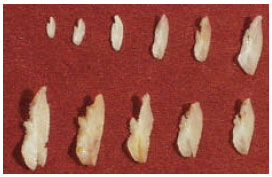 | |
| Fig. 5: | Differences in shape and size according to the different length classes and age groups of sagittae of Caranx caninus (right sagitta, external aspect), increased 2 times |
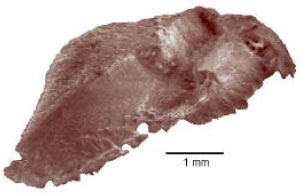 | |
| Fig. 6: | Scanning photograph of the left sagitta, internal aspect of Caranx caninus showing the acoustic canal |
 | |
| Fig. 7: | Scanning photograph on the base of the acoustic canal in the left sagitta, internal aspect of Caranx caninus, showing the calcic carbonate crystals |
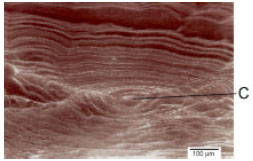 | |
| Fig. 8: | Core (C) and growth rings of the right sagitta, external aspect of Caranx caninus |
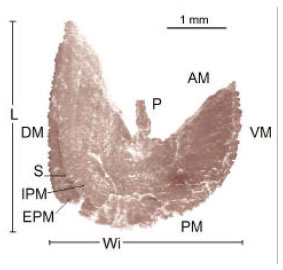 | |
| Fig. 9: | Scanning photograph of the left asteriscus, internal aspect of Caranx caninus, showing its characteristics: AM = anterior margin, PM = posterior margin, P = projection, DM = dorsal margin, VM = ventral margin, L = length, Wi = width, S = sulcus, IPM = internal posterior margin and EPM = external posterior margin |
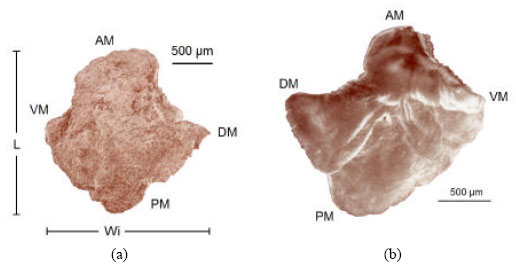 | |
| Fig. 10: | Scanning photograph of the right lapillus of Caranx caninus, a) external aspect, b) internal aspect, showing its main characteristics: AM = anterior margin, PM = posterior margin, DM = dorsal margin, VM = ventral margin, L = length and Wi = width. |
The average width of the sagitta is contained 3.13 times in the average length.
The posterior border is curvilinear and is run lengthwise by the sulcus which divides the otolith in two bodies; the first one with a larger radio is the external aspect (external posterior border), the one with a smaller radio is the internal aspect of the otolith (internal posterior margin) (Fig. 9), which is in contact with the macula and with the neuromast cells (Fig. 3).
The dorsal margin is rectilinear from its anterior to the medium and posterior section of the asteriscus. The ventral margin is curvilinear from the anterior to the posterior margin of this structure. The external aspect is convex and its border is dented, the internal aspect is concave and its border is smooth.
The shape of the asterisci can vary from an individual to another, but these differences are not as large as in the case of the sagittae.
Description of the Lapillus
The anterior margin of this otolith is curvilinear, with a pointed triangular form at the dorsal side and a ventral blunt margin. Both margins narrow towards the rear section and transform into a spherical structure thicker than the rest of the otolith; it exhibits serrated borders formed by crystal growth disposed in epitaxial form.
The external aspect is convex and its surface can be smooth or serrated (Fig. 10a). The internal aspect is concave and has several lobes (Fig. 10b). The otolith curvature increases as the fish grows; and in both aspects of the lapillus the growth patterns of the crystals exhibit different shapes and sizes.
Morphometric Analysis of the Pacific Jack Caranx caninus
The calculated sample sizes are: for sagitta, 31 individuals; asteriscus, 38 and lapillus, 39. There were no morphometric differences between the otoliths of male and female specimens.
Growth of the Sagitta
Table 1 shows the measurements of the sagitta for different length classes. Table 2 expresses the relationship between length and width of the sagitta: the exponent b = 0.791 corresponds to a negative allometric growth; the determination index of R2 = 0.893 and F = 981 of the ANOVA, indicate that the sagitta grows more in width than in length. Also, the allometric relationship between the length of the rostrum and of the antirostrum is expressed in the growth allometric index b = 0.950, with the highest values of R2 and F obtained for this analysis, i.e., 0.964 and 3132, respectively. The tendency of this index is isometric, which shows that among the sizes and age groups, the length of the antirostrum decreases very slowly and disappears in old ages whence it remains as a protuberance (Fig. 4 and 5).
The slope of the relationship between the length and weight of the sagitta is b = 2.595 (R2 = 0.936 and F = 3001) which indicates a negative allometric growth, meaning that as the fish increases its length and age the deposit of protein and calcic carbonate decrease in the sagitta; this phenomenon is probably related with a different orientation of the metabolic products that go to the formation of sexual products and fatty acid storage instead of fish length growth (Gallardo-Cabello, 1986a). This indicates that the growth of otoliths is eccentric in different size classes and ages of fishes, i.e., the postrostrum grows more than the rostrum and the antirostrum; likewise, the dorsal margin grows more than the ventral margin and finally, the deposit of material is higher in the internal than in the external aspect.
| Table 1: | Rostrum (RL) and antirostrum (AL) length, width (Wi) and weight (We) in different size classes of the sagitta of C. caninus |
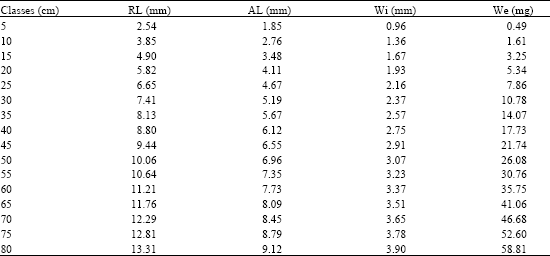 | |
| Table 2: | Relationships between the rostrum and other measures of the sagitta |
 | |
| Table 3: | Relationships between the standard length of the fish (SL) and the measures of the sagitta |
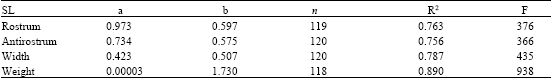 | |
| Table 4: | Length (L), width (Wi) and weight (We) in different size classes of the asteriscus of C. caninus |
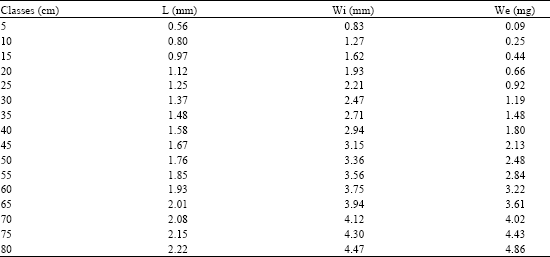 | |
Table 3 shows the relationship between the fish length and the length, width and weight of the sagitta. The highest value of the allometric index, b = 0.597, relates the standard length of the fish with the rostrum, this means that there is a proportionality between the increase of the length of the sagitta and of the fish. The growth in length of this organism can be described by this structure. A high significance of the correlation between the analyzed structures was found by the high values of the determination index R2 and anova (F). When relating very small structures (in millimeters) with values of the corporal length of the fish (in centimeters), the allometric growth indexes tend to be lower than 1.
Growth of the Asteriscus
Table 4 shows the measures of the asteriscus. The relationship between the length and the width (Fig. 9) is expressed by the allometric index b = 1.077 (R2 = 0.855 and F = 661), which is an isometric growth wherein the increment in length is proportional to the width; it means that the dorsal margin grows in proportion to the otolith surface (Table 5).
The relationship between length and weight of the asteriscus is a negative allometric growth index of b = 2.478 (R2 = 0.597 and F = 166), due to the characteristics of this structure which shows considerable thinness, a large surface and a small volume.
According to this, the asteriscus growth is eccentric with respect to the core, which means: that the anterior and dorsal margin grows more than the posterior and ventral margin.
The relationship between fish length and otolith length, its width and weight is shown in Table 6. The most accurate relationship is between the fish length and asteriscus length according to R2 and F. Proportionality between the growth of the asteriscus and of the fish, is showed by the values of the allometric index 0.494 and 0.606, which indicates that, it is possible to determine the age groups through these structures.
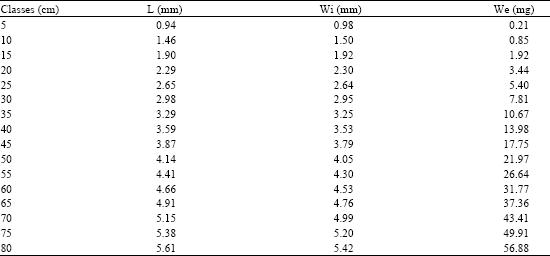
Growth of the Lapillus
The results of the relations of the lapillus are shown in Table 7 (Fig. 10). There is a high tendency to isometric growth index between the length and width of the lapillus (Table 8): b = 0.949 (R2 = 0.981 and F = 6255) which indicates a proportionality growth between length and width of this structure.
The relationship between the length and the weight of the lapillus (Table 8) also shows an isometric growth, b = 3.047 (R2 = 0.971 and F = 3927).
The analyses indicate that the growth of the lapillus is eccentric with respect to the core; it grows more at the anterior and dorsal margin than at the posterior and ventral margin. In the anterior and the posterior margin a larger amount of material is settled for which reason its thickness decreases considerably as fishes grow.
Table 9 shows the relationships between fish length and the length, width and weight of the lapillus. The high value of R2 and F (0.873 and 810 respectively) and the slope b = 0.646 show a high proportionality of the growth of the lapillus and that of the fish, which makes this otolith adequate for age determination.
Identification of the Growth Rings
Six age groups were identified through the analysis of the growth rings in the otolith sagittae (Table 10). One hundred percent of the sagittae otoliths showed perfectly defined growth rings. The area where the deposit of the material takes place with more intensity and the growth rings can be observed more clearly is from the middle half of the dorsal margin to the postrostrum of the otolith (Fig. 4).
| Table 5: | Relationships between the length and the other measures of the asteriscus |
| Table 6: | Relationships between the standard length of the fish (SL) and the measures of the asteriscus. |
 | |
| Table 7: | Length (L), width (Wi) and weight (We) in different size classes of the lapillus of C. caninus |
 | |
| Table 8: | Relationships between the length and the other measures of the lapillus |
| Table 9: | Relationships between the standard length of the fish (SL) and the measures of the lapillus |
 | |
| Table 10: | Number of rings and average length (cm) of C. caninus observed in the sagittae and asterisci |
 | |
The asterisci showed a similar number of rings as the sagittae, only in 25 percent of the cases; it was not possible to identify growth rings in 75 % of the otoliths.
Torres-Aguilar (2002) reports more age groups than those found in this study, organisms until age ten. Nevertheless, the average length for each age is smaller, showing organisms of 54.57 cm in the 6 age group, instead of 58.75 cm in Colima. This reduction in the mean length for each age in the individuals of Michoacán and Guerrero could be related to the phenomenon exposed by Taylor (1958, 1960), who explains that organisms in higher latitudes tend to have a bigger average length at the same age than those in lower latitudes.
It was not possible to identify growth rings in the lapilli, because of the thickness of the structure and the ornamentations of the surface of the external aspect, which avoids the observation of growth rings with transmitted light. Perhaps marks could be easily observed in very thin layered cuts of the lapilli. The irregular deposit of the material on the surface of the external aspect of the lapillus, as well as the high diversity in the orientation patterns of the crystal axes could be related with the piezoelectric capacity of the otoliths for the impulse transmission.
Future studies on the physiology of the otoliths will make the structure and the function of the asterisci and the lapilli more comprehensible.
ACKNOWLEDGMENTS
We owe Dr. Jorge Espino-Vela special thanks for his encouragement on publishing this study and providing helpful comments and suggestions on the manuscript and to Dra. Mariel Haydeé for the corrections of the English version. M. S. Jacqueline Cañetas, was a very helpful technician in the scanning microscope at the UNAM and Dr. René Marquez-Millán with Figure 5. We always thank the fishermen who kindly proportionate the study specimens and from whom we always learn something new on each species we study.
REFERENCES
- Begg, G.A., S.E. Campana, A.J. Fowler and I.M. Suthers, 2005. Otolith research and application: Current directions in innovation and implementation. Mar. Freshwater Res., 56: 477-483.
Direct Link - Berg, E., T.H. Sarvas, A. Harbitz, S.E. Fevolden and A.B. Salberg, 2005. Accuracy and precision in stock separation of North-East Arctic and Norwegian coastal cod by otoliths-comparing readings, image analyses and a genetic method. Mar. Freshwater Res., 56: 753-762.
Direct Link - Bolz, G.R. and R.G. Lough, 1988. Growth through the first six months of Atlantic cod, Gadus morhua and haddock, Melanogrammus aeglefinus, based on daily otolith increments. Fish. Bull., 86: 223-236.
Direct Link - Campana, S.E. and J.D. Neilson, 1985. Microstructure of fish otoliths. Can. J. Fish. Aquat. Sci., 42: 1014-1032.
CrossRefDirect Link - Carlstrom, D., 1963. A crystallographic study of vertebrate otoliths. Biol. Bull., 125: 441-463.
Direct Link - Degens, E.T., W.G. Deuser and R.L. Haedrich, 1969. Molecular structure and composition of fish otoliths. Mar. Biol., 2: 105-113.
CrossRef - Dannevig, E.H., 1956. Chemical composition of the zones in cod otholiths. J. Cons., 21: 156-159.
CrossRef - Francis, C., S.J. Harley, S.E. Campana and P. Doering-Arjes, 2005. Use of otolith weight in length-mediated estimation of proportions at age. Mar. Freshwater Res., 56: 735-743.
Direct Link - Hickling, C.F., 1931. The structure of the otolith of the hake. Q. J. Microsc. Sci., 74: 547-562.
Direct Link - Holst, E., H. Kaiser, L. Schoen, G. Roebig und G. Goldner, 1950. Otolith functioning in fish. Review of comparative Physiology. (Die Arbeitsweise de Statolithenapparates bei Fischen. Zeitschrift fur Verglaichende Physiologie, 32: 60-120.
CrossRef - Lagardere, F., 1989. Influence of feeding conditions and temperature on the growth rate and otolith-increment deposition of larval Dover sole (Solea solea) (L.). Rapp. P. Bv. Reun. Cons. Int. Explor. Mer., 191: 390-399.
Direct Link - Pannella, G., 1971. Fish Otoliths: Daily growth layers and periodical patterns. Science, 173: 1124-1127.
Direct Link - Solomon, G., K. Matsushita, M. Shimizu and Y. Nose, 1985. ge and growth of rose bitterling in Shin Tone river. Bull. Jap. Soc. Sci. Fish., 51: 55-62.
Direct Link - Suthers, I.M., K.T. Frank and S.E. Campana, 1989. Spatial comparison of recent growth in postlarval Atlantic cod (Gadus morhua) off southwestern Nova Scotia: Inferior growth in a presumed nursery area. Can. J. Fish. Aquat. Sci., 46: S113-S124.
CrossRefDirect Link - Taylor, C.C., 1960. Temperature, growth and mortality-the Pacific cockle. J. Conseil., 26: 117-124.
CrossRef - Victor, B.C. and E.B. Brothers, 1982. Age and growth of the fallfish Semotilus corporalis with daily otolith increments as a method of annulus verification. Can. J. Zool., 60: 2543-2550.
CrossRef