
Orathai Wangsantitham
School of Science, University of Phayao, Phayao 56000, Thailand
Paweena Pothong
School of Science, University of Phayao, Phayao 56000, Thailand
Jatuporn Tungjitwitayakul
School of Science, University of Phayao, Phayao 56000, Thailand
Nujira Tatun
School of Science, University of Phayao, Phayao 56000, Thailand
LiveDNA: 66.37684
Journal of Entomology
Year: 2022 | Volume: 19 | Issue: 1 | Page No.: 20-29







ABSTRACT
Background and Objective: Termites have been classified into morphology-physiologically distinct castes that have a broad range of diets. In this study, the candidate of wood-feeding and fungus-growing termites was chosen to examine the relation between gut morphology, the density of microorganisms and starch hydrolyzing enzyme. Materials and Methods: The gut morphologies of the soldiers of wood-feeding (Globitermes sulphureus (Haviland, 1898), Microcerotermes crassus (Snyder, 1934) and fungus-growing (Macrotermes annandalei (Silvestri, 1914) and Odontotermes feae (Wassman, 1896) termites were determined and compared with those of the workers. Results: No significant differences were observed in the morphological structures between the soldiers and workers in different species. However, the microbial cell counts in the guts of the fungus-growing termites were 3-fold higher than those in the wood-feeding termites, with the hindgut having the highest number of microbes. In the wood-feeding termites, the workers and soldiers harboured a similar number of microorganisms, whereas, the workers of the fungus-feeding termites had 3-fold higher levels of microbes than the soldiers. The enzymatic activity of α-amylase in the fungus-growing termites was dramatically higher than that in the wood-feeding termites. However, there were no differences in enzyme activity in the gut segments of the wood-feeding group and the fungus-growing group. Conclusion: Taken together, the different feeding behaviour and castes of the termites affected the density of symbionts in their gut and α-amylase enzymatic activity was modified according to the amount of nutrition in their diets. This study provided additional information on the hidden role of microorganisms in starch metabolism in the termite’s gut.
PDF Abstract XML References Citation
Copyright: © 2022. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Orathai Wangsantitham, Paweena Pothong, Jatuporn Tungjitwitayakul and Nujira Tatun, 2022. Fungus-feeding, alimentary canal, α-amylase, gut, symbiont, termite, cell counts, starch metabolism. Journal of Entomology, 19: 20-29.
DOI: 10.3923/je.2022.20.29
URL: https://scialert.net/abstract/?doi=je.2022.20.29
DOI: 10.3923/je.2022.20.29
URL: https://scialert.net/abstract/?doi=je.2022.20.29
INTRODUCTION
Termites contribute substantially to carbon and nitrogen turnover in many ecosystems, especially in tropical forests. The diets of termites consist mainly of cellulose in various stages of decomposition ranging from wood to humus1. The higher termites have a broad dietary range that includes soil, humus, leaf litter, grass and fungal material. Termites are classified as social insects and each caste shows various behaviours and plays different roles in the colony. Worker, soldier, reproductive and immature castes cooperate in an integrated manner in the termite colony. Most of the termite soldiers have elongated and well-developed mandibles, which play a defensive role in the colony2. However, they are unable to feed themselves; thus, workers are responsible for the transfer of nutritional material from foraging workers to soldiers, meaning the soldiers rely entirely on workers for nutrition. It has been reported that there are differences in gut morphology among worker termites with different feeding behaviors3. Therefore, additional observations of gut morphology in the soldier caste, which relies on the workers for food, would provide further information for understanding the digestive processes of termites.
Thus far, it has been recognized that an association with microbial symbionts in the alimentary canal plays a key role in the digestive process of termites. Since termites express different feeding behaviours, microbial composition in termite’s alimentary canal may have appeared differently. The density and distribution of microorganisms in the alimentary canal of the soil-feeding termite Procubitermes aburiensis (Sjöstedt, 1926) showed that the rectum had 4-fold the microbial density in the crop. The number of bacterial cells in the wood-feeding termite Nasutitermes corniger (Motschulsky, 1855) also varies among the gut compartments4.
In addition to cellulose, plants also contain other polysaccharides, including starch and it has been reported that termites can hydrolyze starch molecules through the action of amylases. The α-amylases are enzymes that hydrolyze α-1,4-glycosidic bonds in amylose chains to produce oligosaccharides, which are then hydrolyzed to glucose by α-glucosidase. Gut extracts from the N. corniger termite can hydrolyze starch molecules in vitro5. Termite α-amylases are reportedly produced in the salivary gland, but they may also have a microbial origin, such as the amylases produced by symbiotic microorganisms in the termite6. However, information on the α-amylases in termites is still limited. In this study, it is observed, the gut morphology of termites of the soldier caste in comparison to those in the worker caste of two species of wood-feeding termites (Globitermes sulphureus (Haviland, 1898) and Microcerotermes crassus (Snyder, 1934): Termitinae) and two species of fungus-growing termites (Macrotermes annandalei) (Silvestri, 1914) and Odontotermes feae (Wassman, 1896): Macrotermitinae). The density of microorganisms in each gut section was examined in both the worker and soldier castes. In addition, the enzymatic activity of α-amylase in the different gut sections and between the soldier and worker castes of the wood-feeding and fungus-growing termites was examined to determine if there is a relationship between microorganism density and α-amylase distribution along the alimentary canal. Moreover, we examined the enzymatic activity of α-amylase in the sponge-like structure of the nesting material inside the mounds of the wood-feeding and fungus-growing termites. These data may reveal a relationship between gut morphology, microbial community and α-amylase, which hydrolyzes starches such as bamboo culm, grass, wood, bark and a certain amount of root7,8 in the termites’ diets, thus allowing them to obtain nutrition from oligo- and monosaccharides as well as from cellulose via hydrolysis. Accordingly, the present study aimed to reveal the relation of the gut morphology, density of microorganisms and the enzymatic activity of α-amylase in termites with different feeding behaviours.
MATERIALS AND METHODS
Insects: The termites G. sulphureus, Mi. crassus, Ma. annandalei and O. feae were collected from a dry dipterocarp forest at the University of Phayao in Phayao, Thailand in 2021. Two castes, including worker and soldier, were collected to examine the gut morphology. The external morphology of the soldier caste was used for termite identification based on the Quick Guide to the Identification of Termites in Thailand by the Forest Research and Development office of the Royal Forest Department.
Termite gut morphology: The external morphology of the worker and soldier termites of G. sulphureus, Mi. crassus, Ma. annandalei and O. feae was observed and recorded by stereomicroscope (Olympus SZX7). Specimens were dissected with fine forceps to observe their alimentary canal and images were captured digitally. The lengths of the whole insect body, whole gut, foregut, midgut and hindgut were measured using Optika Vision Lite 2.1 software.
Microorganisms in the termite gut: The whole guts of workers and soldiers were placed on concave slides containing 100 μL of 20 mM phosphate buffer and dissected under a stereomicroscope into the foregut, midgut and hindgut (mixed segment and first proctodeal, paunch, colon and rectum). Gut samples were squeezed to release their content and the diluted gut content was transferred to 0.2 mL microcentrifuge tubes. More specifically, the hindguts of the wood-feeding termites G. sulphureus and Mi. crassus was separated into proctodeal segments, including the mixed segment (P1+P2), paunch (P3), colon (P4) and rectum (P5). The number of microorganisms in each sample were counted using a hemocytometer under a phase-contrast microscope. The microorganism densities in the gut sections of the four species of termites were compared.
Measuring α-amylase activity: Specimens of the four species of worker termites (G. sulphureus, Mi. crassus, Ma. annandalei and O. feae) were collected and used for the α-amylase activity assays. Soldier specimens were dissected to collect gut samples, which were separated into the foregut, midgut and hindgut. The samples were homogenized in 20 mM sodium acetate buffer at pH 5.5 containing 10 mM NaCl and 20 mM CaCl2. The homogenate was sonicated three times for 5 sec each to break up the cells. Then the homogenate was filtrated through a triple layer of cheesecloth followed by centrifugation at 10,000×g for 15 min at 4°C to remove small debris. The clear supernatant was added to a plastic microcentrifuge tube and stored at -20°C for the α-amylase activity assays. In addition, nest material from both soldier and worker termites of the four species was collected and prepared for enzyme extraction and the α-amylase activity assay as described above. Before the α-amylase assay, the measurement of protein amount in each sample was carried out using a protein dye-binding method (Bio-Rad, Hercules, CA, USA). The reaction mixture contains 5 μL of termite extract and 100 μL of starch solution (0.2% starch, Sigma, St. Louis, MO, USA) in 20 mM sodium phosphate buffer at pH 6.0. The reaction tube was incubated at 37°C for 10 min, then the enzymatic reaction was stopped by the addition of 20 μL of 1 M HCl. The iodine solution (0.5% I2 and 5% KI, 100 μL) was added to the reaction tube and was then transferred to a 1.5 mL cuvette. The amount of starch remaining in the reaction was measured at 580 nm by UV-Vis spectrophotometer (Biomate 3S Spectrophotometer, Thermo Fischer Scientific Inc., MA, USA). The α-amylase activity is expressed as μg of digested starch μg/protein/hr. Three replicates were examined for each experiment.
Statistical analysis: One-way Analysis of Variance (ANOVA) and a least-significance-difference (LSD) multiple-range tests were used for all statistical analyses (SPSS program version 11.5). The significance level was set to 0.05 (p<0.05).
RESULTS
Body length of termites and their gut morphology (Fig. 1a-p): The workers of G. sulphureus (Fig. 1a), Mi. crassus (Fig. 1b), Ma. annandalei (Fig. 1c) and O. feae (Fig. 1d) had body lengths of 4.89, 3.77, 7.17 and 3.75 mm, respectively. While the soldiers had body lengths of 5.01, 4.20, 8.95 and 5.65 mm, respectively (Fig. 1i-l). The gut of these termites were generally divided into three main sections foregut, midgut and hindgut (Fig. 1e-h and m-p).
The termite guts were divided into 3 parts, the foregut, midgut and hindgut. The length of each gut segment was measured in both worker and soldier castes (Table 1). The foregut is a small short tube with a salivary gland (grape-like sac) at the anterior and a crop next to the oesophagus. The foreguts of the workers and soldiers of the wood-feeding termites G. sulphureus and Mi. crassus was 1.88±0.05, 2.46±0.01, 1.68±0.81 and 1.24±0.02 mm in length, respectively. The midguts had a narrow tubular shape that was longer than the foreguts, 3.08±0.05, 3.16±0.02, 2.57±0.11 and 2.84±0.13 mm, respectively. Malpighian tubules, long small tubular shapes attached to the gut, were present at the junctions to the midgut and hindgut. The hindguts were 8.30±0.07, 7.12±0.09, 8.20±0.17 and 6.01±0.01 mm long, respectively. At the anterior part of the hindgut was a narrow tube called the mixed segment. The paunch was an enlarged spherical shaped sac at the anterior and cone-shaped at the posterior. This was followed by the colon and rectum.
In the worker and soldier fungus-growing termites Ma. annandalei and O. feae, the foreguts were similar to those of the wood-feeding termites, 2.45±0.04, 3.37±0.02, 0.99±0.01 and 2.76±0.03 mm long, respectively, while the midguts were 7.98±0.03, 6.10±0.01, 2.84±0.05 and 1.81±0.09 mm long (Table 1). The anterior portion of the midguts exhibited a narrow tubular shape, while the posterior portion was enlarged with a curved shape. The hindguts of the fungus-growing termites were not as complicated as those of the wood-feeding termites, having a tubular shape similar to the midgut, a paunch that was not prominent, followed by the colon and rectum. The hindguts were 6.41±0.15, 6.00±0.04, 3.66±0.05 and 2.64±0.10 mm long, respectively.
 |
| Fig. 1(a-p): | Photographic presentation of the external morphology and intestinal tract of worker (upper panel) and soldier (lower panel) castes of Globitermes sulphureus, Microcerotermes crassus, Macrotermes annandalei and Odontotermes feae Specimen contains the head part at the far left and the intestinal tract is divided into foregut (f), midgut (m) and hindgut (h), scale bar = 1 mm, the paunch in the hindgut part is indicated by a red arrowhead |
| Table 1: | Length of foregut, midgut and hindgut of wood-feeding termite (Globitermes sulphureus and Microcerotermes crassus) and fungus-growing termite (Macrotermes annandalei and Odontotermes feae) | |||
Gut length (mm) | ||||
| Scientific names | Caste | Foregut | Midgut | Hindgut |
| Globitermes sulphureus | Worker | 1.88±0.05 | 3.08±0.05 | 8.30±0.07 |
Soldier | 2.46±0.01 | 3.16±0.02 | 7.12±0.09 | |
| Microcerotermes crassus | Worker | 1.68±0.81 | 2.57±0.11 | 8.20±0.17 |
Soldier | 1.24±0.02 | 2.84±0.13 | 6.01±0.01 | |
| Macrotermes annandalei | Worker | 2.45±0.04 | 7.98±0.03 | 6.41±0.15 |
Soldier | 3.37±0.02 | 6.10±0.01 | 6.00±0.04 | |
| Odontotermes feae | Worker | 0.99±0.01 | 2.84±0.05 | 3.66±0.05 |
Soldier | 2.76±0.03 | 1.81±0.09 | 2.64±0.10 | |
Microbial cell counts in the termite gut: It is examined the number of microorganisms in the guts of the 4 different species of termites (Fig. 2). In the wood-feeding termite G. sulphureus, the paunch of the hindgut (P3) had the highest number of microorganisms (30.78±8.63×106 cells mL–1) in comparison with the other gut sections, the foregut, midgut, P1+P2, P4 and P5 had 7.67±1.65, 3.40±0.34, 6.65±3.37, 2.83±1.15 and 10.73±8.31×106 cells mL–1, respectively (p<0.05, Fig. 2a). Similarly, the Mi. crassus (wood-feeding termite) paunch (P3) had the highest number of microorganisms (40.56±18.39×106 cells mL–1) relative to the other gut sections, with the foregut, midgut, P1+P2, P4 and P5 having 8.41±2.93, 9.18±4.33, 13.57±1.24, 17.43±9.88 and 12.76±4.60×106 cells mL–1, respectively (p<0.05, Fig. 2b). In contrast, in the fungus-growing termite Ma. annandalei, the hindgut had the highest number of microorganisms (109.04±25.51×106 cells mL–1) relative to the other gut sections, including the foregut and midgut, which had 5.64±1.32 and 28.43±5.25×106 cells mL–1, respectively (p<0.05, Fig. 2c).
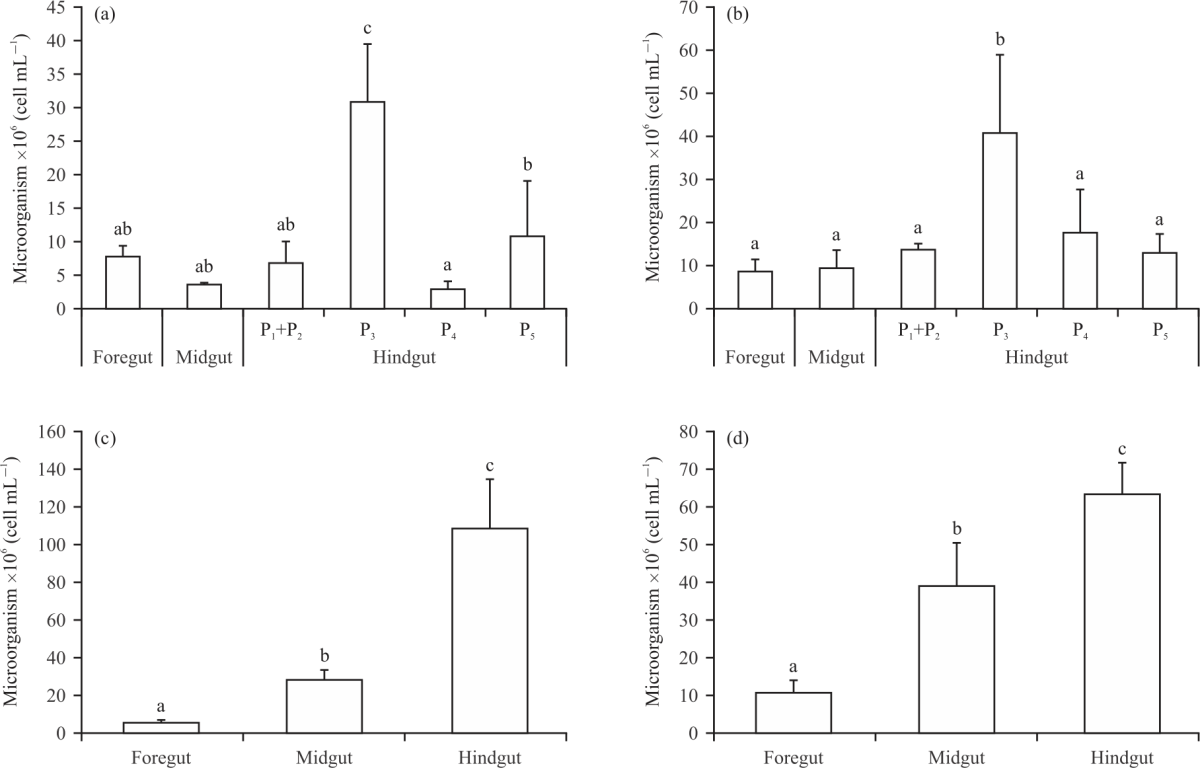 |
| Fig. 2(a-d): | Microbial cell count in different parts of the intestinal tract of (a) Globitermes sulphureus, (b) Microcerotermes crassus, (c) Macrotermes annandalei and (d) Odontotermes feae workers Hindgut was separated into 5 proctodeal segments, error bars indicate SD, the values labelled with different letters are significantly different (p<0.05) |
Similarly, the hindgut of O. feae had the highest number of microorganisms (63.40±8.34×106 cells mL–1) compared to the other gut sections, with the foregut and midgut having 10.50±3.41 and 39.33±11.02×106 cells mL–1, respectively (p<0.05, Fig. 2d).
It is also examined the total number of microorganisms along with the guts of the 4 different species and found no differences between the worker and soldier castes of the wood-feeding termites G. sulphureus and Mi. crassus (Fig. 3a, b). The total numbers of microorganisms in the G. sulphureus workers and soldiers were 40.63±0.26 and 48.04±7.80×106 cells mL–1 (p<0.05, Fig. 3a), respectively, while those in Mi. crassus were 39.82±10.79 and 44.60±4.46×106 cells mL–1, respectively (p<0.05, Fig. 3b). Interestingly, the numbers of microorganisms in the fungus-growing termites (Ma. annandalei and O. feae) were higher in the workers than in the soldiers. In Ma. annandalei, the total numbers of microorganisms in the worker and soldier castes were 119.64±10.55 and 35.56±4.13×106 cells mL–1, respectively (p<0.05, Fig. 3c), while in O. feae (worker and soldier), there were 59.73±1.05 and 16.24±3.74×106 cells mL–1, respectively (p<0.05, Fig. 3d).
α-amylase activity in the wood-feeding and fungus-growing termites: It is measured that, the α-amylase activity in the whole bodies of the wood-feeding and fungus-growing termites (Fig. 4). In the mature workers of G. sulphureus, Mi. crassus, Ma. annandalei and O. feae, the α-amylase enzyme activities were 3.81±0.19, 5.51±0.56, 58.35±0.56 and 51.34±1.30 μg digested starch μg/protein/hr, respectively.
It is also measured that, the α-amylase activity in the foregut, midgut and hindgut of mature soldiers (Fig. 5). For both species of wood-feeding termites (G. sulphureus and Mi. crassus), a low level of activity was detected in all three gut sections. The enzymatic activities in the foregut, midgut and hindgut of G. sulphureus were 4.29±0.49, 4.68±0.64 and 4.52±0.25 μg digested starch μg/protein/hr, respectively (Fig. 5a-c). Similarly, Mi. crassus exhibited enzymatic activities of 5.51±0.56, 4.48±0.12 and 4.28±0.78 μg digested starch μg/protein/hr, respectively (Fig. 5a-c).
In contrast, the α-amylase activity in the fungus-growing termites was demonstrably higher than that in the wood-feeding termites.
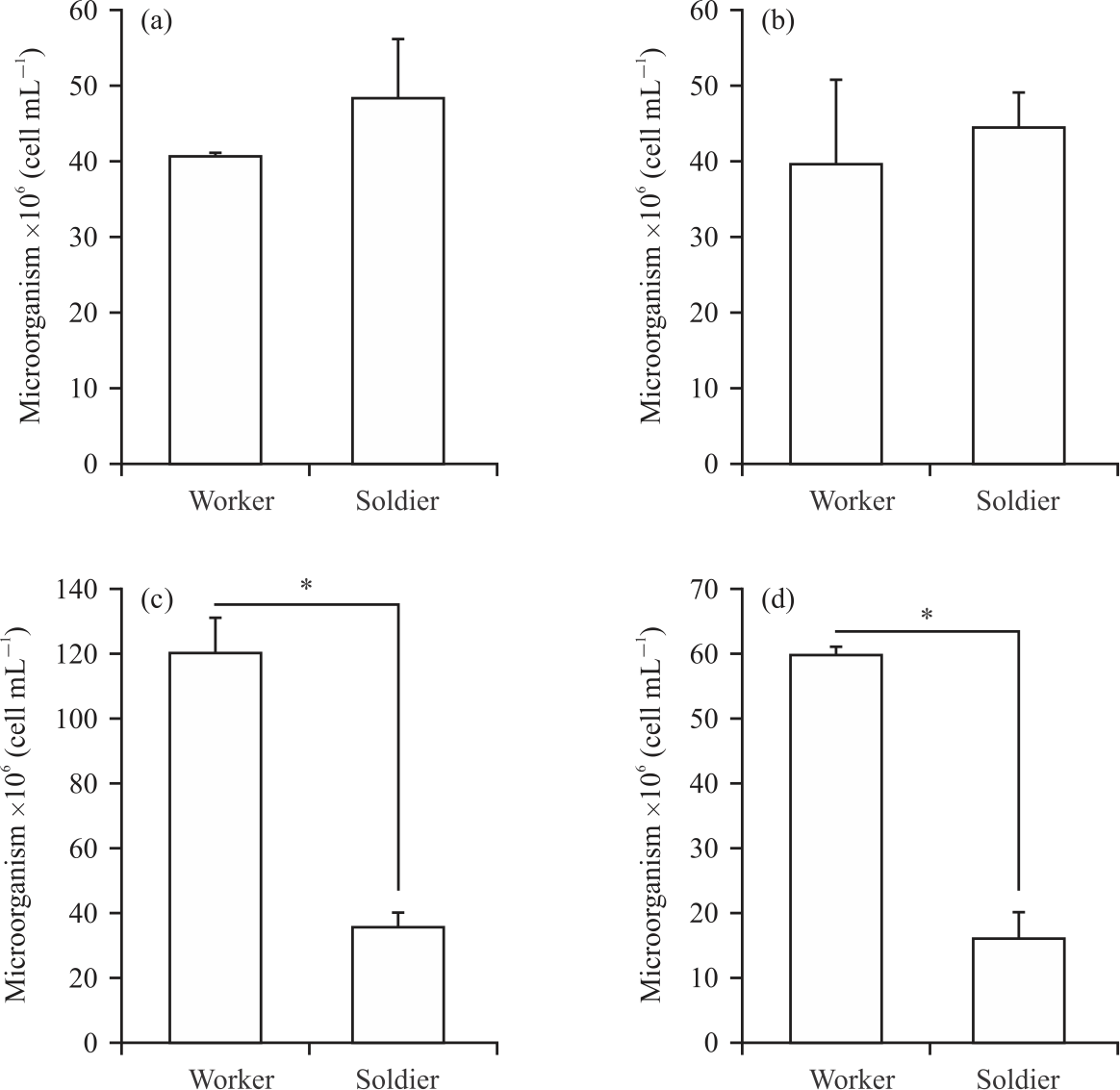 |
| Fig. 3(a-d): | Comparison of microbial cell count in the intestinal tract of workers and soldiers in (a) Globitermes sulphureus, (b) Microcerotermes crassus, (c) Macrotermes annandalei and (d) Odontotermes feae workers Error bars indicate SD, the asterisks above the bars indicate a significant difference between the soldier and worker castes within species (p<0.05) |
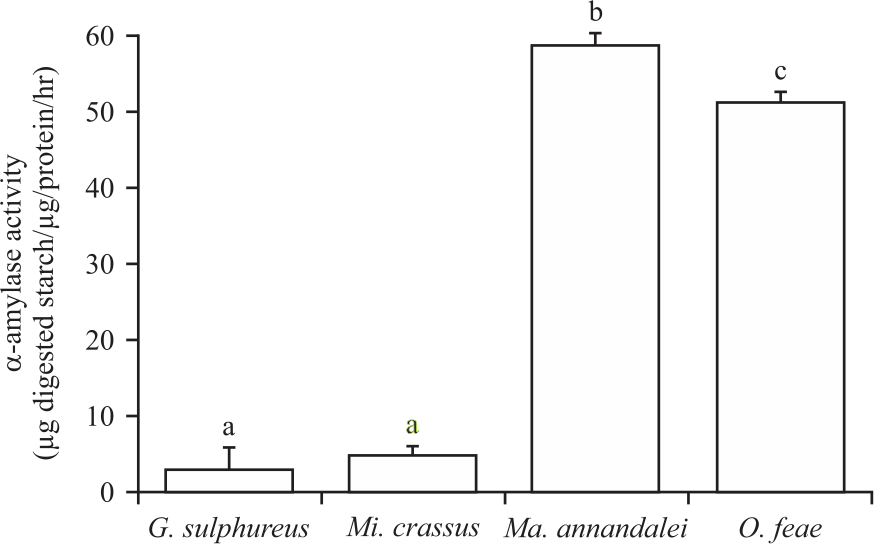 |
| Fig. 4: | Enzymatic activity of α-amylase in Globitermes sulphureus, Microcerotermes crassus, Macrotermes annandalei and Odontotermes feae workers Enzymatic activity of α-amylase is expressed as μg digested starch μg/protein/hr, error bars indicate SD, the values labelled with different letters are significantly different (p<0.05) |
In Ma. annandalei, the enzymatic activities were 175.00±10.22, 230.45±20.1 and 140.05±11.62 μg digested starch μg/protein/hr in the foregut, midgut and hindgut, respectively, while in O. feae, they were 156.12±4.95, 196.00±2.96 and 139.42±13.59 μg digested starch μg/protein/hr, respectively (Fig. 5a-c).
Exogenous α-amylase activity: In addition, It is examined, the α-amylase activity in the nest materials (Fig. 6). Very low levels of α-amylase activity were found in the nest materials of the wood-feeding termites G. sulphureus and Mi. crassus, 5.19±0.65 and 5.38±1.21 μg digested starch μg/protein/hr, respectively. Interestingly, high levels of α-amylase activity were found in the nest materials (fungus comb) of the fungus-growing termite’s Ma. annandalei and O. feae, 351.77±20.41 and 350.89±11.01 μg digested starch μg/protein/hr, respectively.
|
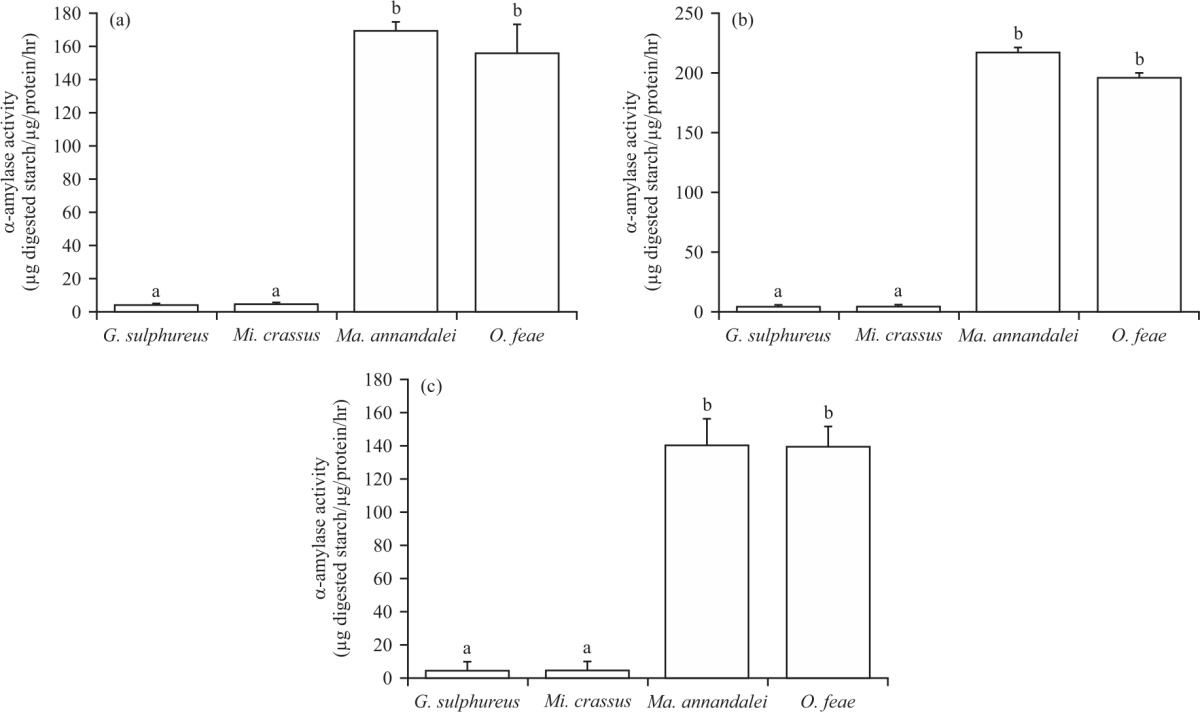
| Fig. 5(a-c): | Distribution of α-amylase activity in the (a) Foregut, (b) Midgut and (c) Hindgut of Globitermes sulphureus, Microcerotermes crassus, Macrotermes annandalei and Odontotermes feae Enzymatic activity of α-amylase is expressed as μg digested starch μg/protein/hr, error bars indicate SD, the values labelled with different letters are significantly different (p<0.05) |
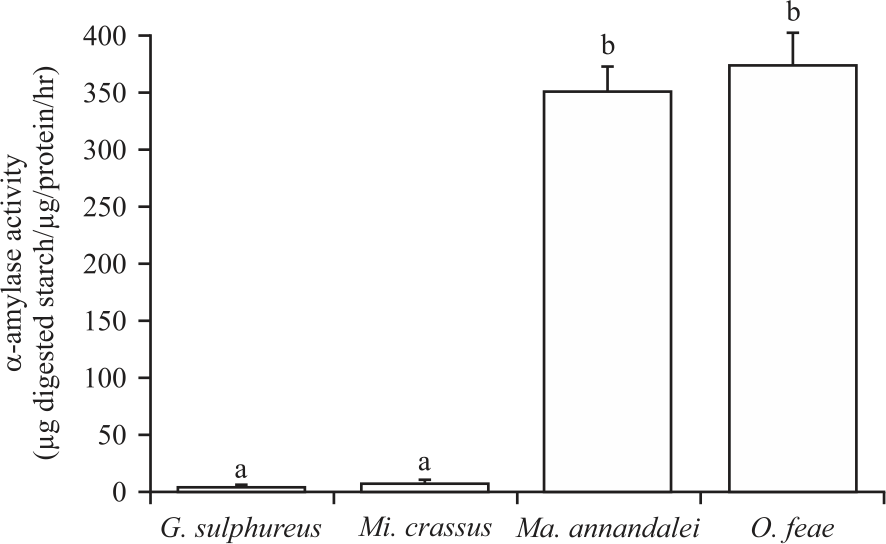 |
| Fig. 6: | Detection of α-amylase activity in nesting material collected from termite’s mound of Globitermes sulphureus, Microcerotermes crassus, Macrotermes annandalei and Odontotermes feae Enzymatic activity of α-amylase is expressed as μg digested starch μg/protein/hr, error bars indicate SD, the values labelled with different letters are significantly different (p<0.05) |
DISCUSSION
The different roles in a colony lead to worker and soldier termites having distinct morphological differences in their body size, colour, head shape and especially mouthparts. Termite soldiers have elongated and well-developed mandibles, which are used for a defensive role in the colony. These modified mouthparts cause soldier termites to be unable to feed themselves and workers are responsible for transferring predigested diets to the soldiers, indicating they rely entirely on workers for nutrition9. Differences in gut morphology have been reported among worker termites with different feeding behaviours, i.e., wood-feeding and fungus-growing termites3. To provide further information for understanding the digestive processes of termites, in this study, we made additional observations of the gut morphology of the soldiers that rely on food from workers. It is found the worker and soldier castes of both wood-feeding and fungus-growing termites had similar morphologies in the foregut, midgut and hindgut, which was long and highly modified. As described by Noirot10, the hindgut of wood-feeding termites is composed of the ileum (P1), enteric valve (P2), paunch (P3), colon (P4) and rectum (P5). More specifically, the diets present in the alimentary canals of the soldiers and workers differed, with what appeared to be a fine particulate in the soldier's gut, indicating consumption of a predigested diet that had been transferred from a worker’s digestive system through trophallaxis behavior11. The length of the hindgut in the wood-feeding termites (G. sulphureus and Mi. crassus) was about 2-fold longer than the midgut and foregut, whereas, in the fungus-growing termites (Ma. annandalei and O. feae), the midgut and hindgut were similar in length. The ratio between gut and body length for G. sulphureus, Mi. crassus, Ma. annandalei and O. feae workers and found the guts of the wood-feeding termites were 2.7 and 3.3-fold longer than their body lengths, whereas, in the fungus-growing termites, they were 2.3 and 1.99-fold longer. The ratios of the gut to body length in the soldier castes were 2.54 and 2.40 in the wood-feeding termites, while those in the fungus-growing termites were 1.72 and 1.32. These ratios indicate the fungus-growing termites had shorter alimentary canals and less modification than those in the wood-feeding termites. Based on phylogenetic studies, the digestive system of termites changes significantly during the evolution of the family Termitidae, which is recorded as one of the evolutionary lineages in termites12. All the termites studied were members of the family Termitidae and belonged to the subfamilies Termitidae and Macrotermitinae. The greatest variation in intestinal morphology is found in the Termitidae family. The flagellated microorganism in the hindgut of termites belonging to the Termitidae family has not been found, suggesting the evolution of the digestive system in both morphological and physiological aspects13.
The diversity of microorganisms inhabiting the guts of various termite feeding classes has been reported as high as 106–107 cells per μL of gut content volume extracted from the whole gut14. More specifically, the microbial cell count in this study showed the paunches of the hindguts in G. sulphureus and Mi. crassus workers had the highest number of cells (3.07×107 and 4.05×107) relative to other gut sections. A study of bacterial diversity in the wood-feeding termite Nasutitermes corniger (Motschulsky, 1855) also reported that the P3 compartment (paunch) of the hindgut contained the highest number of microbial cells (1.5×107), 2-fold higher than the other gut sections5. Interestingly, the number of microbial cells in the fungus-growing termites in this study, Ma. annandalei and O. feae, was about 2.5-fold higher than that in the wood-feeding termites examined. Moreover, since they have distinctive feeding behaviours, we compared the number of microbial cells in the guts of worker and soldier termites. Interestingly, the worker and soldier wood-feeding termites (G. sulphureus and Mi. crassus) harboured similar numbers of microbial cells in their whole gut, which could be explained by the trophallaxis behaviour of the workers orally transferring pre-digested food containing gut microorganisms to the soldier termites. In contrast, the worker castes of the fungus-growing termites harboured 3-fold more microbial cells in their guts than the soldier caste.
The evolution of termites involves the presence of intestinal microorganisms, including bacteria, Archaea and protists15,16. Fungus-growing termite workers (subfamily Macrotermitinae), including genera Macrotermes and Odontotermes, mix plant material with the asexual spores of their plant-degrading fungal symbiont (Termitomyces) in their guts and deposit the predigested plant as a sponge-like structure called a fungus comb inside the mound. Young worker termites consume the mycelia and later, older fungus comb is consumed by the termites17. This could explain why the microbial cell counts in Ma. annandalei and O. feae were so high in the workers because their guts were used as inoculation sites for the fungal symbiont. This was supported by observations that microorganisms in the guts of fungus-growing termites are accompanied by a large number of fungal spores. The microbial symbionts in the hindgut are horizontally transferred to nestmates such as soldiers and other reproductive castes in the colony via proctodeal feeding18.
Because plant materials often contain both amylose and cellulose, which are linear glucose-based polymers distinguished by the glycosidic linkage between the glucose units, the microorganisms in termite guts are thought to have amylolytic activity. It has been reported that α-amylases are one of the important enzymes involved in growth and longevity during larval and adult stages in insects belonging to the order of Orthoptera, Hymenoptera, Diptera, Lepidoptera and Coleoptera7. In the termite Nasutitermes walker (Hill, 1942), α-amylase was found mainly in the midgut (73%), followed by salivary glands (18%) and also found in other gut portion19. In contrast, α-amylase activity was at a high level in salivary glands in Mastotermes darwiniensis (Froggatt, 1897) (lower termite), which was about 81% of α-amylase activity in the whole body9. In this study, α-amylase was high in the worker castes of the fungus-growing termites Ma. annandalei and O. feae, while the wood-feeding termites had less α-amylase activity. Hence, amylase derived from the fungus-growing termite guts could provide a novel alternative source for various factory applications, as amylase is an enzyme that plays an important role in the industry, with amylase production comprising about 25% of the enzyme trade20. It has been reported that insect α-amylase activity is affected by different types of diets. In Helicoverpa armigera (Hübner, 1808), which regulates α-amylase enzyme levels in response to the macromolecular composition of their diet, α-amylase activity is high when fed a diet containing a high concentration of carbohydrates and low when fed plants with a low concentration of carbohydrates21. Termite food, such as leaves, grass, wood, living plants and especially the fungi fed by fungus-growing termites, generally contains a relatively large amount of starch9. Macrotermes gilvus (Hagen, 1858) has been reported to have higher amylase activity than wood-feeding termites (Coptotermes formosanus (Shiraki, 1909) and Reticulitermes speratus (Kolbe, 1885)), digesting at least 7 μg of starch when consuming leaf litter. The α-amylases in fungus-growing and wood-feeding termites can have two origins: First, that produced by the termite tissues and second, that produced by microorganisms in the termite’s gut content. Metagenomic analysis has revealed that there are numerous microorganisms in the guts of the wood-feeding termites Nasutitermes ephratae (Holmgren, 1910) and N. corniger and some of the groups analyzed were able to produce α-amylase (glycoside hydrolase 1322). Moreover, bacteria with amylolytic activity has been reported in the guts of the wood-feeding termite Coptotermes sp.23. This study found high α-amylase activity in the fungus comb of fungus-growing termites and at a lower activity level, in the nest material of wood-feeding termites. This was following a report that termitarium collected from Macrotermes bellicosus (Smeathman, 1781) exhibited α-amylase activity, with enzymatic activity levels higher than that in the salivary glands24. The activity level is suspected to be due to contributions by the microorganisms present in the termitarium soil. However, the α-amylase activity in the termitarium may also be derived from the saliva of the many worker termites involved in building the mound.
In any case, the degree that which symbionts contribute to the hydrolysis of starch in the intestines of termites remains unclear. The RNA interference of the termite amylase gene (from termite cells) may be able to provide information on how much microbial amylase (in gut content) contributes to starch hydrolysis in termites. Thus, it remains to be clarified whether the microorganisms in termites’ guts contribute to their starch digestive capacity to fulfill their energy demands.
CONCLUSION
In conclusion, the structure of the gut in wood-feeding and fungus-growing termites were similar, but the density of microorganisms is present throughout the gut and high in the hindgut portion. The difference in gut microorganism density between soldier and worker castes was found in fungus-growing termites only. The enzymatic activity of α-amylase was recorded throughout the gut, however, the hindgut had the highest activity of α-amylase. In addition, α-amylase was also detected in nest material, indicating that α-amylase may be originated from both termite cells and microorganisms.
ACKNOWLEDGMENT
This work was supported by the revenue budget in 2020, School of Science, University of Phayao (PBTSC63015) awarded to O.W.
REFERENCES
- Rust, M.K. and N.Y. Su, 2012. Managing social insects of urban importance. Annu. Rev. Entomol., 57: 355-375.
CrossRefDirect Link - Koshikawa, S., T. Matsumoto and T. Miura, 2002. Morphometric changes during soldier differentiation of the damp-wood termite Hodotermopsis japonica (Isoptera, Termopsidae). Insectes Soc., 49: 245-250.
CrossRefDirect Link - Tatun, N., C. Sawatnathi, S. Tansay and J. Tungjitwitayakul, 2017. Comparison of gut morphology and distribution of trehalase activity in the gut of wood-feeding and fungus-growing termites (Isoptera: Termitidae). Eur. J. Entomol., 114: 508-516.
CrossRefDirect Link - Köhler, T., C. Dietrich, R.H. Scheffrahn and A. Brune, 2012. High-resolution analysis of gut environment and bacterial microbiota reveals functional compartmentation of the gut in wood-feeding higher termites (Nasutitermes spp.). Appl. Environ. Microbiol., 78: 4691-4701.
CrossRefDirect Link - de Albuquerque Lima, T., E.V. Pontual, L.P. Dornelles, P.K. Amorim and R.A. Sá et al., 2014. Digestive enzymes from workers and soldiers of termite Nasutitermes corniger. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 176: 1-8.
CrossRefDirect Link - Subekti, N. and T. Yoshimura, 2009. α-amylase activities of saliva from three subterranean termites: Macrotermes gilvus Hagen, Coptotermes formosanus Shiraki, and Reticulitermes speratus (Kolbe). Jpn. J. Environ. Entomol. Zool., 20: 191-194.
CrossRefDirect Link - Okahisa, Y., T. Yoshimura and Y. Imamura, 2006. Seasonal and height-dependent fluctuation of starch and free glucose contents in moso bamboo (Phyllostachys pubescens) and its relation to attack by termites and decay fungi. J. Wood Sci., 52: 445-451.
CrossRefDirect Link - Qamar, S.F.A., S.M. Cunningham and J.J. Volenec, 2006. Phosphate nutrition and defoliation effects on growth and root physiology of alfalfa. J. Plant Nutr., 29: 1387-1403.
CrossRefDirect Link - Ishikawa, Y., H. Aonuma and T. Miura, 2008. Soldier-specific modification of the mandibular motor neurons in termites. PLoS ONE, Vol. 3.
CrossRefDirect Link - Noirot, C., 2001. The gut of termites comparative anatomy, systematics, phylogeny II - Higher termites. Ann. Soc. Entomol. Fr., 37: 431-471.
Direct Link - Nalepa, C.A. 2015. Origin of termite eusociality: Trophallaxis integrates the social, nutritional, and microbial environments. Ecol. Entomol., 40: 323-335.
CrossRefDirect Link - Inward, D.J.G., A.P. Vogler and P. Eggleton, 2007. A comprehensive phylogenetic analysis of termites (Isoptera) illuminates key aspects of their evolutionary biology. Mol. Phylogenet. Evol., 44: 953-967.
CrossRefDirect Link - Bignell, D.E., Y. Roisin and N. Lo, 2011. Biology of Termites: A Modern Synthesis. 2nd Edn., Springer, Dordrecht, ISBN: 978-90-481-3976-7, Pages: 576.
CrossRefDirect Link - Brauman, A., J. Doré, P. Eggleton, D. Bignell, J.A. Breznak and M.D. Kane, 2006. Molecular phylogenetic profiling of prokaryotic communities in guts of termites with different feeding habits. FEMS Microbiol. Ecol., 35: 27-36.
CrossRefDirect Link - Abe, T., D.E. Bignell and M. Higashi, 2000. Termites: Evolution, Sociality, Symbioses, Ecology. 1st Edn., Springer, Netherlands, ISBN: 978-0-7923-6361-3, Pages: 466.
CrossRefDirect Link - Eggleton, P., 2000. Global Patterns of Termite Diversity. In: Termites: Evolution, Sociality, Symbioses, Ecology, Abe, T., D.E. Bignell and M. Higasi (Eds)., Springer, Dordrecht, ISBN: 978-90-481-5476-0, pp: 25-51.
CrossRefDirect Link - Wisselink, M., D.K. Aanen and A. van 't Padje, 2020. The longevity of colonies of fungus-growing termites and the stability of the symbiosis. Insects, Vol. 11.
CrossRefDirect Link - Ohkuma, M. and A. Brune, 2011. Diversity, Structure, and Evolution of the Termite Gut Microbial Community. In: Biology of Termites: A Modern Synthesis, Bignell, D.E., Y. Roisin and N. Lo (Eds.), Springer, Dordrecht, ISBN: 978-90-481-3976-7, pp: 413-438.
CrossRefDirect Link - Hogan, M., P.C. Veivers, M. Slaytor and R.T. Czolij, 1988. The site of cellulose breakdown in higher termites (Nasutitermes walkeri and Nasutitermes exitiosus). J. Insect Physiol., 34: 891-899.
CrossRefDirect Link - Reddy, N.S., A. Nimmagadda and K.R.S.S. Rao, 2003. An overview of the microbial α-amylase family. Afr. J. Biotechnol., 2: 645-648.
Direct Link - Kotkar, H.M., P.J. Sarate, V.A. Tamhane, V.S. Gupta and A.P. Giri, 2009. Responses of midgut amylases of Helicoverpa armigera to feeding on various host plants. J. Insect Physiol., 55: 663-670.
CrossRefDirect Link - Warnecke, F., P. Luginbuhl, N. Ivanova, M. Ghassemian and T.H. Richardson et al., 2007. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature, 450: 560-565.
CrossRefPubMedDirect Link - Mulyani, P.D., R.M. Hamid, R.Z. Janatunaim and Y.A. Purwestri, 2018. Amylolytic ability of bacteria isolated from termite (Coptotermes sp.) gut. Indones. J. Biotechnol. 23: 14-20.
CrossRefDirect Link - Popoola, K.O.K. and A.V. Opayele, 2012. Morphometrics of Macrotermes bellicosus (African mound termite) (Blattodea: Termitidae) and the impact of its saliva amylase on the strength of termitarium soil. N. Y. Sci. J., 5: 207-216.
Direct Link