
Lamharchi El Hassane
Laboratory of Biotechnology and Valorization of Natural Resources, Faculty of Sciences, Ibn Zohr University, B.P. 8106, Agadir, Morocco
Zahidi AbdelAziz
Laboratory of Biotechnology and Valorization of Natural Resources, Faculty of Sciences, Ibn Zohr University, B.P. 8106, Agadir, Morocco
LiveDNA: 212.31502
Hallouti Ayoub
Laboratory of Microbial Biotechnology and Plant Protection, Faculty of Sciences, Ibn Zohr University, B.P. 8106, Agadir, Morocco
Akchour Abdellah
Laboratory of Biotechnology and Valorization of Natural Resources, Faculty of Sciences, Ibn Zohr University, B.P. 8106, Agadir, Morocco
El Finti aissam
Laboratory of Biotechnology and Valorization of Natural Resources, Faculty of Sciences, Ibn Zohr University, B.P. 8106, Agadir, Morocco
Boubaker Hassan
Laboratory of Microbial Biotechnology and Plant Protection, Faculty of Sciences, Ibn Zohr University, B.P. 8106, Agadir, Morocco
LiveDNA: 212.8065
El Mousadik Abdelhamid
Laboratory of Biotechnology and Valorization of Natural Resources, Faculty of Sciences, Ibn Zohr University, B.P. 8106, Agadir, Morocco
LiveDNA: 212.23672
Journal of Entomology
Year: 2020 | Volume: 17 | Issue: 2 | Page No.: 48-57







ABSTRACT
Background and Objective: The California Red Scale (CRS) is a serious pest of citrus in many parts of the world. Different control methods have been adopted, but their effectiveness remains limited. The objective was to isolate fungi from the CRS corpses and determine the efficacy of two isolated strains, Aspergilllus flavus and Fusarium oxysporum against different CRS developmental stages. Materials and Methods: Fungal isolates obtained from RSC corpses were identified by studying their macroscopic and microscopic aspects. Mycelial biomass was used for genomic DNA extraction according to kit protocol. Polymerase Chain Reaction (PCR) with specific primers on a partial sequence was used for each fungal isolate. Each PCR product was sequenced in the forward and reverse directions using an Applied Biosystem ABI 3130×l sequencer. Results: The result of the study informs that most isolated strains belonged to five genera of Aspergillus, Alternaria, Fusarium, Rhizopus and Cladosporium. At concentration of 106 conidia mL–1, pre-nymph male and adult male under shield showed greater mortality ranging from 84.02-93%, but for female stages, this rate did not exceed 58%. Adult males, male pre-pupae and mobile larvae (68.3-100%) are more affected by Fusarium than Aspergillus strain. Conclusion: It can be concluded that the two entomopathogenic fungi tested can be considered as good candidates for biological control of California Red Scale (CRS). Therefore, they will be used as microbial agents in Integrated Pest Management (IPM) strategies.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Lamharchi El Hassane, Zahidi AbdelAziz, Hallouti Ayoub, Akchour Abdellah, El Finti aissam, Boubaker Hassan and El Mousadik Abdelhamid, 2020. Biological Control of Aonidiella aurantii (Hemiptera-Diaspididae) by the Entomopathogenic Fungi Aspergilllus flavus and Fusarium oxysporum. Journal of Entomology, 17: 48-57.
DOI: 10.3923/je.2020.48.57
URL: https://scialert.net/abstract/?doi=je.2020.48.57
DOI: 10.3923/je.2020.48.57
URL: https://scialert.net/abstract/?doi=je.2020.48.57
INTRODUCTION
Aonidiella aurantii (Mask.) (Hemiptera: Diaspididae) is a cosmopolitan and polyphagous insect which is considered as one of the most important pests of citrus in California, Australia, Mediterranean basin as well in Morocco1-3. This pest can infest all the above ground parts of citrus trees by sucking the sap on the plant tissue with their long filamentous mouth parts, causing thus deformations by the toxic action of saliva. In addition to an encrusting on fruits, the sole presence of scales on fruits considerably reduces their market value causing huge economic losses since these infested fruits can be down graded in the packing stations4-6. Because of the great economic importance of damages caused by California Red Scale (CRS), producers had resorted to chemical control for several years especially against spring generation which causes fruit damages. The main method of control of CRS estimated difficult and is frequently followed by re-current infestations in a short period, in addition to elimination of natural enemies present in the field. Despite, several alternative methods to chemical control currently seems to be used in conventional orchards, such as mating disruption, mass trapping as well as cultural methods, but damages remain considerable7-10. In Morocco, from 1966, the ectoparasitoid Aphytis melinus was introduced and used to regulate CRS populations. Other native or introduced parasitoids or predators have also been used to control CRS populations such as Aphytis chrysomphali, A. lingnanensis, Encarsia citrine, Comperiella bifasciata as well as Coleoptera such as Exochomus nigromaculatus, E. nigropictus and E. quadripustulatus 11-14. Despite this specific richness, some works was carried out in Souss (more than 34% of Moroccan citrus production), only the two species A. melinus and C. bifasciata contribute effectively to the reduction of A. aurantii populations in conventional orchards15.
The improvement of Integrated Pest Management (IPM), especially against CRS requires monitoring of populations dynamic of this pest and knowledge of the natural enemy species composition in each climatic zone. To enhance this control, attention is focusing on Entomopathogenic Fungi (EPF) which are considered as an effective biological alternative to insecticides. Metarhizium and Beauveria belongs to Clavicipitaceae and Cordycipitaceae, respectively. Both fungal genera are insect pathogens and are ubiquitous in soils world-wide16-18. One technique not yet tested in California red scale, is the use of entomopathogenic fungi, which has produced good results with some Hemiptera insects19. Some authors have exposed pests to suspensions of fungal strains directly or simultaneously with predators. The aims of this study were to isolate fungi from the CRS corpses, identify molecularly and determine the efficacy of two isolated strains, Aspergilllus flavus and Fusarium oxysporum against different developmental stages of this pest in order to use these microbial agents in IPM programs.
MATERIALS AND METHODS
The study was carried out at the two laboratories Biotechnologies and Valorization of Natural Resources and Microbial Biotechnologies and Plant Protection of Faculty of Sciences, Ibn Zohr University, Agadir, Morocco from January, 2018-April, 2019. Molecular analysis (DNA extraction and sequencing) were carried out at the Technical Support Units (TSU) for scientific research at the National Center for Scientific and Technical Research (CNRST), Rabat, Morocco.
Collection of insect corpses and fungi isolation: The corpses of red californian scale were collected in a biological farm in Ain Chaib, Ouled Teima (30°24'N, 9°13'W) located between Agadir and Taroudant in Souss plain in the south west of Morocco. Samples were collected from infested trees; collection of dead insects was performed on twigs, leaves and fruits. These corpses once isolated, are kept in a petri dish at ambient temperature. They were disinfected with bleach at 0.1% for 2 min to remove microbial flora. These corpses are then rinsed several times with distilled water according to the method proposed by Jouda et al.20. Subsequently, corpses were deposited in petri dishes containing the Potato Dextrose Agar (PDA) medium and incubated at temperature of 25±2°C until, the significant multiplication of the fungus. Purification of the isolates is carried out, following successive subcultures of each fungus by the removal of fungal explants present at the borders of colonies grown on PDA medium.
Identification of fungal isolates: Fungal isolates obtained were identified by studying their macroscopic and microscopic aspects using identification keys21,22. The macroscopic aspect concerned color, diameter and back of colony. Microscopic aspect concerned mycelium morphology and conidia by observation of mycelium fragments and fruiting forms.
Pathogenicity test: Among fungi isolated from CRS corpses, some isolates of both genera Aspergillus and Fusarium showed signs of virulence in laboratory conditions. Aspergillus spp. is well known as a saprophyte rather than an entomopathogen fungus, it was unclear if the fungus was responsible for pest mortality. To establish this, some infested fruits are treated with fungal conidia.
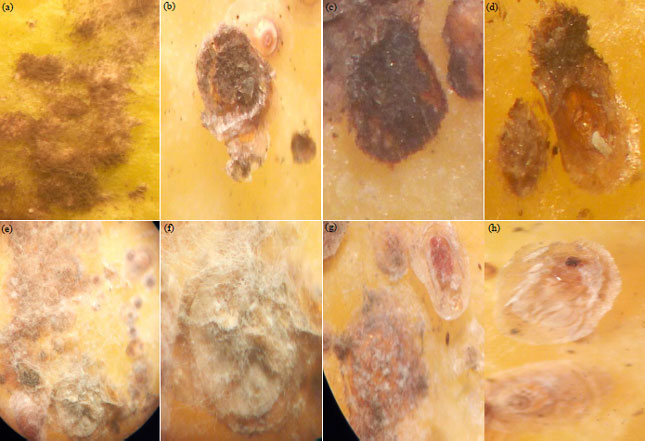 | |
| Fig. 1(a-h): | Symptomatology of fungal mycosis caused by Aspergillus flavus and Fusarium oxysporum on the California red scale (Aonidiella aurantii) at the stage of dissemination of the two fungi with dense filamentous masses (a) Infested instars of CRS on fruit, (b) L2 stage infested, (c) F1 female infested, (d) Adult male under shield infested, (e) Infested instars of CRS on fruit , (f) F1 female infested, (g) Infested male pre-nymph, (h) Adult male under shield infested |
The treated insects showed symptoms of mycosis within 48 h of treatment (Fig. 1). Thus, 2 isolates Fusarium and Aspergillus were selected for an in vivo test against california red scale. After 2 weeks of incubation at 25°C, spore suspension was prepared. The conidia were harvested directly by scraping surface of the culture medium, followed by homogeneous suspension in sterile distilled water containing 0.02% Tween 80 (Tween 80 is used to facilitate the conidia separation). The concentration of suspension was determined by using a Malassez cell, then diluted with distilled water to produce conidial concentrations. For each strain, several spores concentration were tested, 105, 106 and 107. According to Hallouti et al.23, the concentration 106 conidia mL–1 was the most effective against Ceratitis capitata. After inocula preparation, infested fruits were inoculated by spraying 2 mL of conidia suspension23,24. Three replicates were performed for each fungal isolate. Sterile distilled water containing 0.02% of Tween 80 was used as control test. Treated fruit with EPF were incubated at 25°C, mortality rates were counted after 24, 48, 72 and 96 h and so until the 9th day in order to determine Lethal Time 50 (LT50)25. A re-isolation of insect entomopathogens has been done to verify that the fungi tested are responsible for mortalities (Koch's postulates). Due to natural mortality in the control fruits (MRc), corrected percentage of mortality in each test (MR) under laboratory conditions was calculated using Abbott’s formula26,27:
Where:
| MR | = | Percentage of mortality observed in different stages of CRS |
| MRt | = | Percentage of CRS mortality in treatment |
| MRc | = | Percentage of mortality in the control test |
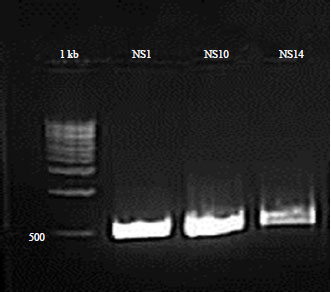 | |
| Fig. 2: | Full-ITS PCR products amplified from Fusarium spp., Aspergillus spp. and Beauveria with ITS1/ITS4 primers, molecular weight markers (100 bp DNA ladder, Promega) |
Extraction of genomic DNA: The fungal cultures isolated in the previous assays were grown on PDA medium for 7 days at 25°C. Mycelial biomass was extracted for DNA analyses according to procedure28 with some modifications by using extraction buffer (100 mM Tris-HCl, 2% CTAB, 1.4 mM NaCl). DNA quality analysis was performed by UV-visible spectrophotometery by measuring the absorbance (or optical density) of the nucleic acids at 260 and 280 nm wavelength. The quality of the DNA solution was accepted by quantifying A260/A280 absorbance ratio which varied28-30 between 1.8 and 2.0. At the level of Molecular Biology and Functional Genomics platform in the National Center for Scientific and Technical Research (CNRST), Rabat, a second genomic DNA extraction from the same filamentous fungi samples maintained in culture on PDA medium using the “Isolate II Plant DNA kit” BIOLINE according to kit protocol.
Amplification of fungal DNA and sequencing: Polymerase Chain Reaction (PCR) with specific primers on a partial sequence was used to confirm morphological identification of each fungal isolate (Fusarium and Aspergillus). PCR reactions were performed using the Internal Transcribed Spacer region (ITS-1) forward (TCCGTAGGTGAACCTGCGG) and ITS-4 reverse (TCCTCCGCTTATTGATATGC) primers for Aspergillus spp.31. For Fusarium spp., 2 primers were used to amplify specifically the ITS regions. PCR fragments (600 bp) amplified with the nucleotide sequences of these primers: 5’-CAACTCCCAAACC CCTGTGA-3’ and 5’-GCGACGATTACCAGTAACGA-3’31-33.
Polymerase chain reaction was performed in a 25 μL volume reactions with the following concentrations: Containing 150 ng of fungal DNA for each fungal isolate, Taq polymerase buffer (5×), 0.2 mM of each dNTP, 1 μL of each primer (ITS1 10 μM and ITS4 10 μM) and 0.2 μL of Taq polymerase 5U μL–1 (MyTaq DNA polymerase kit, Bioline)32-35. The reaction mixture was supplemented with 2 mM of MgCl2 concentration. The PCR was performed in a thermal cycler (Verity, ABI 3130×L), using a program that start with an initial denaturation cycle at 95°C for 1 min, followed by 35 thermal cycles of denaturation for 15 sec at 95°C, annealing for 20 sec at 57°C followed by extension for 15 sec at 72°C and ending with final extension cycle at 72°C for 3 min. The reaction was held at 4°C. The presence of PCR products was confirmed by electrophoresis for 2 h at a constant voltage (90 V) on agarose gel (1.5%) in the presence of a molecular weight marker of 100 bp. The visualization is carried out by the documentation system "G Box" as shown in Fig. 2. Each PCR product was sequenced in the forward and reverse directions using an Applied Biosystem ABI 3130×L sequencer (Genetic Analyzer, 16 capillaries, Applied Biosystem). Sequences were submitted to Basic Local Alignment Search Tool (BLAST). Internal transcribed spacer 1, partial sequence; 5.8 ribosomal RNA gene and Internal transcribed spacer 4, complete sequence and large sub-unit ribosomal RNA gene, partial sequence were used for Fusarium spp. and Aspergillus spp. identification (Basic Local Alignment Search Tool).
Data analysis: The degree of virulence of the 2 isolates tested was estimated according to the ability of the fungus to induce death of various stages of california red scale. In addition, calculation of Lethal Time 50 (LT50) was also carried out, since at the beginning of experiment there were 50 fruits with 3 repetitions for each fungus tested (Fusarium spp. and Aspergillus spp.) as well as the control test. After every 24 h, 9 fruits (3×3 = 9) are analyzed and eliminated immediately. Analysis of Variances (ANOVA) with two factors (fungus strain and pest stage) in crossed model was adopted. Comparisons were made with Newman-Keuls test using a significance level of 95%. All statistical analyzes were performed using the Statistica Version 6 software.
RESULTS
Identification of fungal isolates: Microscopic identification of fungi collected directly from A. aurantii corpses revealed the presence of mycoflor which is represented by different species and genera such as; Aspergillus niger, Aspergillus (spp.), Alternaria spp., Fusarium spp. 1, Fusarium spp. 2, Rhizopus and Cladosporium (Fig. 3).
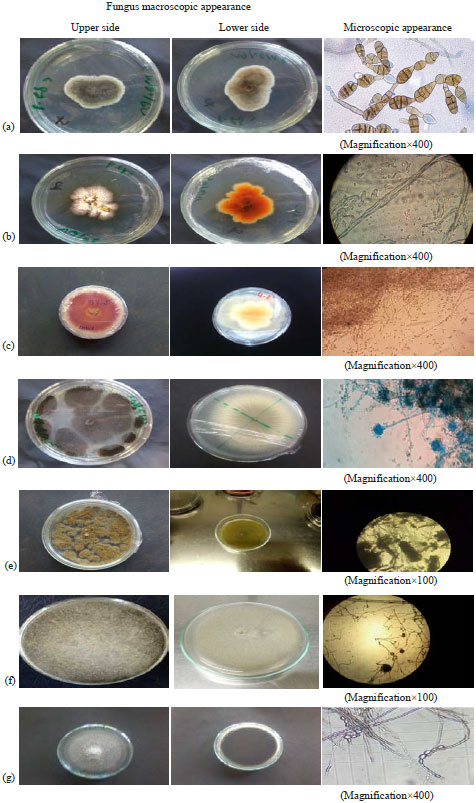 | |
| Fig. 3(a-g): | Fungus macroscopic appearance, microscopic appearance and name of the genus or species isolated from A. aurantii corpses (Mounting liquid: Cotton blue), (a) Alternaria spp., (b) Fusarium spp. 1, (c) Fusarium spp., 2: Fusarium oxysporum, (d) Aspergillus spp. 1: Aspergillus niger, (e) Aspergillus spp. 2: Aspergillus flavus, (f) Rhizopus spp. and (g) Cladosporium spp. |
The majority of these microorganisms are developed saprophytically on insects. Alternaria spp. is characterized by a filamentous thallus, branched and partitioned, conidia are multicellular, isolated with a long beak at the end. The colonies are olive-green in color with a greyish surface, the periphery is slightly irregular in light color.
| Table 1: | Analysis of the variance of mortality rate of california red scale subjected to effects of fungal strains (106 conidia mL–1) |
 | |
| Df: Degree of freedom, SS: Sum of squares, MS: Mean square, ***Highly significant | |
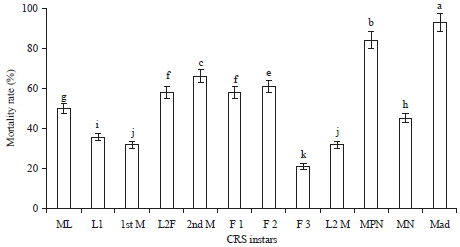 | |
| Fig. 4: | Mean rate of mortality induced by both fungi at different California Red Scale (CRS) instars (male adult under the shield), a, b …. k: Homogeneous groups obtained by means comparison method Least Significant Difference (LSD) at an alpha significance level of 5% |
The color of the underside is dark gray with a clear gray margin and significant sporulation. Aspergillus niger is characterized by colonies formed by white to yellowish compact mycelium covered by a dense layer of conidiophores with variable colors and white margins. The reverse is colorless. The conidiophore is smooth, hyaline or brownish in their upper half. For Aspergillus spp., the aerial mycelium is undeveloped, greenish yellow in color and with a light yellow in the lower side. The conidiophores widen, progressively upwards to form vesicles of variable diameters and are terminated by a fungal vesicle of the genus Aspergillus. Conidia are greenish yellow in color and vary widely in size. Fusarium colonies are generally characterized by an aerial mycelium of greyish-white color with the appearance of a flattened cone. The periphery is often characterized by whitish pigmentation during the first days after isolation, then with a red wine color as well as by an abundance of ellipsoidal microconidia and macroconidia of variable size. Rhizopus spp. has a cottony mycelium that turns brown-black at maturity. Zygospores are round or oval with a brown-black color, suspensory swollen, often unequal. For Cladosporium spp., color of colonies was greenish black and surface was velvety without wrinkles. The reverse of colonies was nearly black. The conidial chains were mostly un-branched and arose directly from the side or the tip of hyphae.
Two isolates showed signs of pathogenesis; these are 2 entomopathogenic fungi belonging to the genera Fusarium and Aspergillus. Inoculation of infested fruits by these 2 strains have allowed the appearance in the 1st day, a fine white down on individuals mummified, which down has grown very rapidly to give dense filamentous masses (Fig. 1). Molecular identification of Aspergillus strain, using ITS1 and ITS4; revealed that ITS amplicons were variable in size from 438 bp for ITS1 to 442 bp for ITS4. Alignment with GenBank reference sequences shows similarity to Aspergillus flavus for both ITS. For the 2nd strain, ITS amplicons from Fusarium was ranged in size from 418 bp for ITS1 to 434 bp for ITS4. Alignment with GenBank sequences shows similarity to Fusarium oxysporum.
Pathogenicity test and lethal time 50 (LT50): Analysis of variance of mortality rate revealed highly significant differences between the 2 fungal strains, as well as CRS instars and developmental stage×fungal strain interaction (Table 1). The sensitivity to both fungal strains was very different according to pest instar. In fact, pre-nymph male (MPN) and adult male under shield (Mad) instars showed greater mortality (Fig. 4). For female stages, the mortality rate did not exceed 62%; the reproductive female was less sensitive to both isolates with a mortality rate less than 25%.
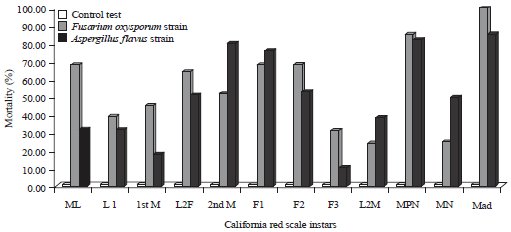 | |
| Fig. 5: | Mortality rates observed per california red scale instars induced by both fungal strains A. flavus and F. oxysporum |
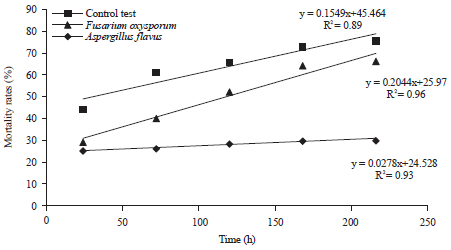 | |
| Fig. 6: | Linear regression of mortality rate of california red scale individuals induced by both fungal strains as a function of incubation time to determine Lethal Time 50 (LT50) |
| R²: Coefficient of determination | |
Younger stages (ML and L1) were less affected. In addition, adult males, male pre-pupae and mobile larvae are more affected by Fusarium oxysporum than Aspergillus flavus. For other instars, Fusarium was responsible for variable mortality reaching around 69% for F1 females. Aspergillus flavus induced mortality especially for second moult, pre-pupae and adult males. The reproductive female (F3) appears to be less affected by the two strains (Fig. 5).
Mean Lethal Time (LT50) defined as time required to induce death of 50% of pest population was also observed. The lowest LT50 was about 29 h and 28 min for Fusarium oxysporum. It was about 117 h and 47 min for Aspergillus flavus strain (Fig. 6). A clear correlation was noted between the mortality rate of the different stages of the pest and the 2 fungal isolates. After more than 200 h, approximately 67% for Aspergillus and 78% for Fusarium of the population exhibit symptoms of pathogenicity (Fig. 1).
DISCUSSION
Entomopathogenic fungi are a broad group of microorganisms, that help to maintain pest abundance below economic levels in several agroecosystems. Limited studies on native isolates of these biological control agents have been done in South West Morocco except some works which seeks to isolate and determine the diversity of the medfly-associated entomopathogenic fungi in Argane forest23. This research endeavors to expand on earlier works and isolate other fungi directly from corpses of another key citrus pest namely Aonidiella aurantii (CRS). The most isolated species belonged to five genera. Fusarium and Aspergillus were more represented. These results are consistent with those obtained by Saiah et al.36 following work on Phyllocnistis citrella or other results on Tuta absoluta (Lepidoptera: Gelechiidae)20,37. These germs in some cases can be developed in saprophytic way on insects. Results on the CRS should be considered as important since works on the armored scale insects are still rare. In Australia, Microcera coccophila has been reported as a parasite of armoured scales such as Aonidiella aurantii and A. citrina38. It is important to determine the factors conditioning the implantation of fungal infection in CRS corpses to control effectively against populations of the scale. Using fungal primers (ITS1 and ITS4), comparison to reference strains demonstrated that both ITS are sufficient for accurate identification of Aspergillus spp. and Fusarium spp. Alignment with reference sequences shows similarity to Aspergillus flavus and Fusarium oxysporum. A number of Fusarium spp. with varying pathogenic abilities against insects are tested at least under laboratory conditions, such as F. semitectum and F. sacchari among different Aphididae species20. Similar result reported that a large number of Fusarium spp. are entomopathogenic, some are weak, facultative pathogens, especially against lepidopteran and coleopteran orders. They will also colonize their dead hosts as saprophytes39. In addition, Fusarium spp. was more virulent against Ceratitis capitata compared to Beauveria and Aspergillus23. It is well known that a high level of intra-species genetic diversity is typical for species undergoing sexual cycle with the meiotic recombination32-40. But using the universal fungal primers (ITS1/ITS4), with high levels of nucleotide diversity provide a high degree of discriminatory power useful to isolate identification as reported in study on Fusarium oxysporum species complex41. This level of diversity is highly observed when aligning isolate strains sequences with reference sequences. In general, F. oxysporum was more virulent than A. flavus mainly against mobile larvae, pre-nymphs and adult males under shield. In addition, all infested insects presented a white down on mummified individuals, which down has increased very rapidly to give dense filamentous masses that develop on corpses. California red scales infected by both strains were easily recognized in the early stages of infection when insects are still alive. The first indication of disease is a slight color change of body, which is normally white, to a pale coral-pink tint. Later, as the disease progresses and the insect dies, the zoosporangia develop internally giving the scale a bright orange color. After the insect is dead, the zoosporangia mature and the insect's body becomes a powdery mass of golden-yellow zoosporangia. If resting zoosporangia are present, all or part of the body may be brick-red in color42. The insect mortality caused by these 2 strains of entomopathogenic fungi may be the result of the following actions: Like action of released toxins, the physical obstruction of circulation, organ invasion and destruction of insect tissue or nutritional deficiency43. California red scale shield provide a structurally and chemically complex barrier to fungus penetration. Generally, fungi can infect insects by direct penetration through the rigid cuticle or in contact with the cuticle. Spores which are considered to be the unit infectious of fungi germinate and penetrate through the integument by combining mechanical and enzymatic pressures on the insect44-46. Differences between F. oxysporum and A. flavus could probably be explained by the mechanisms involved during infestation. Infection process will be similar to other fungal pathogens of insects and includes recognition of host, spores attachment until next round of spore’s production. Young stages and adult males are more vulnerable than adult females. This result will be considered as very important for this pest in this region. In future, CRS population management programs, essentially against spring population which causes significant damages, knowledge of population dynamic is a necessity to program a treatment by these microorganisms in field and improve their efficiencies. Similarly, combination of the 2 strains at the same time in a single product to target different CRS stages would be an alternative to manage this key pest of citrus in Morocco. The implementation of several microbial control agents under difficult environmental conditions (cold weather, dry weather, etc.) can be effective and serve as potential alternatives pest’s management such as CRS, Ceratitis capitata and would appear to be a new track to develop in order to limit the use of broad-spectrum chemical insecticides.
CONCLUSION
To minimize the harmful impact of pests on citrus production, development of new approaches using entomopathogenic fungi would be an effective alternative for controlling CRS. Fusarium oxysporum and Aspergillus flavus can be considered as good candidates for biological control of california red scale. Their use would be a very promising alternative to ensure phytosanitary protection respectful of environment and human health.
SIGNIFICANCE STATEMENT
This study discovers two native strains of entomopathogenic fungi that can be beneficial for biological control of Aonidiella aurantii, responsible of serious damage on fruit production in Morocco. This study will help the researcher to discover other strains as potential candidates in biological control to reduce the use of pesticides that many researchers were not able to explore. Thus, the testing of these two strains on other pests such as phytophagous mites and other dipterans is to be explored.
ACKNOWLEDGMENTS
We gratefully acknowledge the Moroccan Ministry of Scientific Research, the presidency of Ibn Zohr University, Morocco for financial support. We offer our sincere thanks to the managers of Agricultural Domains for the logistical and financial support for the realization of this work. Our thanks to administration, technical service of CNRST (National Center for Scientific and Technical Research) for their technical assistance and molecular analyzes. We thank the anonymous reviewers and office journal which provided helpful comments that greatly improved the manuscript.
REFERENCES
- Guirrou, Z., I. El Kaoutari, A. Boumezzough, M. Chemseddine and A. Hilal, 2003. Contrôle des populations d'Aonidiella aurantii (Maskelle) en vergers d'agrumes au Maroc. Fruits, 58: 3-11.
CrossRefDirect Link - Washington, J.R. and G.P. Walker, 1990. Histological studies of California red scale (Homoptera: Diaspididae) feeding on citrus. Ann. Entomol. Soc. Am., 83: 939-948.
CrossRefDirect Link - Jacas, J.A. and A. Urbaneja, 2010. Biological Control in Citrus in Spain: From Classical to Conservation Biological Control. In: Integrated Management of Arthropod Pests and Insect Borne Diseases, Ciancio, A. and K. Mukerji (Eds.)., Integrated Management of Plant Pests and Diseases, Vol. 5. Springer, Dordrecht, pp: 61-72.
- Rodrigo, E., P. Troncho and F. García-Marí, 1996. Parasitoeds (Hym.: Aphelinidae) of three scale insects (Hom.: Diaspididae) in a citrus grove in Valencia, Spain. Entomophaga, Vol. 41, No. 1.
CrossRefDirect Link - El-Sayed, A., D. Suckling, C. Wearing and J. Byers, 2006. Potential of mass trapping for long-term pest management and eradication of invasive species. J. Econ. Entomol., 99: 1550-1564.
CrossRefPubMedDirect Link - Miller, J.R., L.J. Gut, F.M. De Lame and L.L. Stelinski, 2006. Differentiation of competitive vs. non-competitive mechanisms mediating disruption of moth sexual communication by point sources of sex pheromone (Part I): Theory. J. Chem. Ecol., Vol. 32, No. 10.
CrossRefDirect Link - Byers, J.A., 2007. Simulation of mating disruption and mass trapping with competitive attraction and camouflage. Environ. Entomol., 36: 1328-1338.
CrossRefDirect Link - Sorribas, J., R. Rodríguez and F. García-Marí, 2010. Parasitoid competitive displacement and coexistence in citrus agroecosystems: Linking species distribution with climate. Ecol. Applic., 20: 1101-1113.
CrossRefDirect Link - Pekas, A., A. Aguilar, A. Tena and F. Garcia-Marí, 2010. Influence of host size on parasitism by Aphytis chrysomphali and A. melinus (Hymenoptera: Aphelinidae) in Mediterranean populations of California red scale Aonidiella aurantii (Hemiptera: Diaspididae). Biol. Control, 55: 132-140.
CrossRefDirect Link - Smaili, M.C., M. Abbassi, J.A. Boutaleb and A. Blenzar, 2013. Richesse spécifique des ennemis naturels associés aux vergers d'agrumes au Maroc: Intérêt et implication pour la lutte biologique. EPPO Bull., 43: 155-166.
CrossRefDirect Link - Boyero, J.R., J.M. Vela, E. Wong, C.G. Ripoll, M.J. Verdu, A. Urbaneja and P. Vanaclocha, 2014. Displacement of Aphytis chrysomphali by Aphytis melinus, parasitoids of the California red scale, in the Iberian Peninsula. Spanish J. Agric. Res., 12: 244-251.
Direct Link - Martínez-Blay, V., J. Pérez-Rodríguez, A. Tena and A. Soto, 2018. Density and phenology of the invasive mealybug Delottococcus aberiae on citrus: Implications for integrated pest management. J. Pest Sci., 91: 625-637.
CrossRefDirect Link - Lamharchi, H., A. Akchour, A. Zahidi and A. El Mousadik, 2016. Control of spring generation of Aonidiella aurantii (Hemiptera, Diaspididae) citrus pest in Souss region. Proceedings of the 3rd International American Moroccan Agricultural Sciences Conference, December 13-16, 2016, Ouarzazate, Morocco.
- Sewify, G.H. and S.M. Habeeb, 2001. Biological control of the tick fowl Argas persicargas persicus by the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae. J. Pest Sci., 74: 121-123.
CrossRefDirect Link - Chouvenc, T., N.Y. Su and J.K. Grace, 2011. Fifty years of attempted biological control of termites-analysis of a failure. Biol. Control, 59: 69-82.
CrossRefDirect Link - Carrillo, D., C.A. Dunlap, P.B. Avery, J. Navarrete and R.E. Duncan et al., 2015. Entomopathogenic fungi as biological control agents for the vector of the laurel wilt disease, the redbay ambrosia beetle, Xyleborus glabratus (Coleoptera: Curculionidae). Biol. Control, 81: 44-50.
CrossRefDirect Link - Faria, M. and S.P. Wraight, 2001. Biological control of Bemisia tabaci with fungi. Crop Prot., 20: 767-778.
CrossRefDirect Link - Jouda, G.J., B.M. Naima and B. Halima-Kamel, 2011. Etudes préliminaires sur les champignons entomopathogènes des pucerons de l’artichaut en Tunisie. Faunistic Entomol., 63: 171-181.
Direct Link - Hallouti, A., A. Zahidi, R. Bouharroud, A. El Mousadik, A.A.B. Aoumar and H. Boubaker, 2017. Diversity of entomopathogenic fungi in argane forest soil and their potential to manage Mediterranean fruit fly (Ceratitis capitata). J. Pharm. Pharmacol., 5: 746-754.
CrossRefDirect Link - Er, M.K., H. Tunaz and A. Gökçe, 2007. Pathogenicity of entomopathogenic fungi to Thaumetopoea pityocampa (Schiff.) (Lepidoptera: Thaumatopoeidae) larvae in laboratory conditions. J. Pest Sci., 80: 235-239.
CrossRefDirect Link - Balogun, S.A. and O.E. Fagade, 2004. Entomopathogenic fungi in population of Zonocerus variegatus (L.) in Ibadan, South West, Nigeria. Afr. J. Biotechnol., 3: 382-386.
CrossRefDirect Link - Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Alves, S.B., L.S. Rossi, R.B. Lopes, M.A. Tamai and R.M. Pereira, 2002. Beauveria bassiana yeast phase on agar medium and its pathogenicity against Diatraea saccharalis (Lepidoptera: Crambidae) and Tetranychus urticae (Acari: Tetranychidae). J. Invertebrate Pathol., 81: 70-77.
CrossRefDirect Link - Passone, M.A., L.C. Rosso, A. Ciancio and M. Etcheverry, 2010. Detection and quantification of Aspergillus section Flavi spp. in stored peanuts by real-time PCR of nor-1 gene and effects of storage conditions on aflatoxin production. Int. J. Food Microbiol., 138: 276-281.
CrossRefDirect Link - Melo, S.C.O., C. Pungartnik, J.C.M. Cascardo and M. Brendel, 2006. Rapid and efficient protocol for DNA extraction and molecular identification of the basidiomycete Crinipellis perniciosa. Genet. Mol. Res., 5: 851-855.
Direct Link - Barra, P., L. Rosso, A. Nesci and M. Etcheverry, 2013. Isolation and identification of entomopathogenic fungi and their evaluation against Tribolium confusum, Sitophilus zeamais and Rhyzopertha dominica in stored maize. J. Pest Sci., 86: 217-226.
CrossRefDirect Link - Jayanthi, P.D.K., A. Arthikirubha, K. Vivek, M.A. Ravindra, G. Selvakumar and V. Abraham, 2015. Aspergillus flavus impairs antioxidative enzymes of Sternochetus mangiferae during mycosis. J. Invertebr. Pathol., 124: 73-77.
CrossRefDirect Link - Irzykowska, L., J. Bocianowski, A. Waśkiewicz, Z. Weber and Z. Karolewski et al., 2012. Genetic variation of Fusarium oxysporum isolates forming fumonisin B1 and moniliformin. J. Applied Genet., 53: 237-247.
CrossRefDirect Link - Abd-Elsalam, K.A., I.N. Aly, M.A. Abdel-Satar, M.S. Khalil and J.A. Verreet, 2003. PCR identification of Fusarium genus based on nuclear ribosomal-DNA sequence data. Afr. J. Biotechnol., 2: 82-85.
CrossRefDirect Link - Mouhaddab, J., A.I.T. Naima, H. Achtak, F. Msanda and A. Zahidi et al., 2015. Patterns of genetic diversity and structure at fine scale of an endangered Moroccan endemic tree (Argania spinosa L. Skeels) based on ISSR polymorphism. Not. Bot. Horti Agrobot. Cluj-Napoca, 43: 528-535.
CrossRefDirect Link - Moricca, S., A. Ragazzi, T. Kasuga and K.R. Mitchelson, 1998. Detection of Fusarium oxysporum f.sp. vasinfectum in cotton tissue by polymerase chain reaction. Plant Pathol., 47: 486-494.
CrossRefDirect Link - Saiah, F., B.S. Bendahmane, M.Y. Benkadda, A. Berkani, W. Lakhdari and N. Kolai, 2010. Isolement de champignons entomopathogènes à partir de Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae). Faunistic Entomol., 63: 199-202.
Direct Link - Badaoui, M.I., A. Berkani and B. Lotmani, 2010. Les entomopathogènes autochtones, nouvel espoir dans le contrôle biologique de Tuta absoluta Meyrick 1917 (Lepidoptera: Gelechiidae) en Algérie. Faunistic Entomol., 63: 165-169.
Direct Link - Dao, H.T., G.A.C. Beattie, A.Y. Rossman, L.W. Burgess and P. Holford, 2015. Systematics and biology of two species of Microcera associated with armoured scales on citrus in Australia. Mycol. Progress, Vol. 14, No. 4.
CrossRefDirect Link - Teetor-Barsch, G.H. and D.W. Roberts, 1983. Entomogenous Fusarium species. Mycopathologia, 84: 3-16.
CrossRefPubMedDirect Link - Kerényi, Z., A. Moretti, C. Waalwijk, B. Oláh and L. Hornok, 2004. Mating type sequences in asexually reproducing Fusarium species. Applied Environ. Microbiol., 70: 4419-4423.
CrossRefDirect Link - O’Donnell, K., C. Gueidan, S. Sink, P.R. Johnston and P.W. Crous et al., 2009. A two-locus DNA sequence database for typing plant and human pathogens within the Fusarium oxysporum species complex. Fungal Genet. Biol., 46: 936-948.
CrossRefDirect Link - Fisher, F.E., 1950. Entomogenous fungi attacking scale insects and rust mites on citrus in Florida. J. Econ. Entomol., 43: 305-308.
CrossRefDirect Link - Hajek, A.E. and R.J. St. Leger, 1994. Interactions between fungal pathogens and insect hosts. Annu. Rev. Entomol., 39: 293-322.
CrossRefDirect Link - Clarkson, J.M. and A.K. Charnley, 1996. New insights into the mechanisms of fungal pathogenesis in insects. Trends Microbiol., 4: 197-203.
CrossRefDirect Link