
Ibrahim A. Hamed
Department of Plant Protection, Faculty of Agriculture, Zagazig University, 44511 Zagazig, Egypt
LiveDNA: 20.30428
Refat M. Sherif
Department of Plant Protection, Faculty of Agriculture, Zagazig University, 44511 Zagazig, Egypt
El-Sayed A. El-Sheikh
Department of Plant Protection, Faculty of Agriculture, Zagazig University, 44511 Zagazig, Egypt
LiveDNA: 20.1579
Aly A. Shallaby
Department of Plant Protection, Faculty of Agriculture, Zagazig University, 44511 Zagazig, Egypt
Journal of Entomology
Year: 2019 | Volume: 16 | Issue: 3 | Page No.: 91-97







ABSTRACT
Background and Objective: The house fly, Musca domestica L., has the ability to develop resistance against pyrethroid insecticides in a short exposure time. This study aimed to screen the susceptibility of house fly to lambda-cyhalothrin and determine the activities of metabolic enzymes in insect. Materials and Methods: House fly populations collected from different locations in Sharkia Governorate, Egypt were used. Bioassay experiments were carried out on 3rd larval instars of each population. Activities of metabolic enzymes were determined. Results: Data of bioassay experiments indicated that population collected from Kafr Sakr is 10.6-fold resistant to lambda-cyhalothrin. Other populations collected from San El-Hagar, Abo Kabir, Abo Hammad and Zagazig showed 8, 9.8, 6.9 and 7.5 fold of resistance as compared on LC50 levels. Biochemical assays revealed that San El-Hagar and Kafr Sakr populations have the highest significant activity of esterase, while Zagazig population showed the highest activity in carboxylesterase recording 2.58 fold higher than laboratory strain followed by populations from Kafr Sakr and San El-Hagar. Results of mixed function oxidase were significantly higher in both Zagazig and Abo Hammad populations with 1.3 and 1.3 fold than what recorded in the laboratory strain. Kafr Sakr and Abo Hammad populations showed the highest significant GST activity recording 174.7 and 170 mmol min–1 g–1 b.wt., respectively. Conclusion: The obtained results showed that populations collected from different locations in Sharkia Governorate have low to moderate resistance to lambda-cyhalothrin with the possible role of carboxylesterase. The existing resistance is a warning to carefully use lambda-cyhalothrin in control programs of this pest in the specified locations.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ibrahim A. Hamed, Refat M. Sherif, El-Sayed A. El-Sheikh and Aly A. Shallaby, 2019. Susceptibility and Biochemical Determination of House Fly (Musca domestica L.) to Lambda-cyhalothrin. Journal of Entomology, 16: 91-97.
DOI: 10.3923/je.2019.91.97
URL: https://scialert.net/abstract/?doi=je.2019.91.97
DOI: 10.3923/je.2019.91.97
URL: https://scialert.net/abstract/?doi=je.2019.91.97
INTRODUCTION
The house fly, Musca domestica L., is one of the major public health insect that adversely affects on both human and animal health by carrying more than 100 disease pathogens1. House flies are spreading not only in dairies and livestock facilities, but also in urban cities causing harmful effects for humans and domestic animals such as nuisance, aesthetic damages to different structures and also play a role in the mechanical transmission of pathogens2.
Control methods for urban pests including house flies are depending mainly on insecticides for being fast acting, relatively cheap and convenient to use3. Synthetic insecticides have been used for decades as a principle control strategy for this pest. Pyrethroids are a group of insecticides that showed importance in pest control and used widely in the control of agricultural and health pests. They account for approximately 25% of the world insecticide market4. Lambda-cyhalothrin, a sodium channel modulator, has been used extensively for the management of various insect pests including M. domestica5.
Due to the quick and easy of insecticides application, these methods become the choice of people to face house flies infestations. However, house flies have the ability to become resistant against insecticides used for their management usually in a short period of time6. Extensive applications have resulted in many incidents of resistance to these insecticides with different mechanisms of resistance i.e., metabolic resistance to pyrethroids in insects can be associated with increase in cytochrome P450 activity, increase in general esterase and elevated glutathione S-transferases due to high genes expression, enabling them to overproduce these types of enzymes7,8. The P450-mediated detoxification and sodium channel-mediated target site insensitivity are major mechanisms related to pyrethroid resistance development in house flies9. Increase resistance levels to insecticides in this pest has become a major issue due to the extensive and poorly optimized application of insecticides10. The purpose of this study was to investigate the current status of house fly susceptibility that collected from 5 localities in Sharkia Governorate, Egypt. In addition this study will provide baseline data that will help to define enzymes that might associate in house fly resistance to pyrethroid.
MATERIALS AND METHODS
Insects and rearing: A laboratory strain was established by collecting house fly larvae from fields in Zagazig, Egypt (during summer of 2016) and reared in the laboratory for 25 generations without any exposure to insecticides. A field strain was collected from dairy farms of 5 locations in Sharkia Governorate, Egypt (San El-Hagar, Abo Kabir, Abo Hammad, Kafr Sakr, Zagazig) during summer of 2017 and 2018. About 500 larvae of Musca domestica L., were collected from each location and brought to the laboratory for rearing. The larvae were reared on a medium containing of powdered milk, sugar, yeast and wheat bran at a ratio of 1.5:1.5:5:20 (w/w), respectively and made a paste with 65 mL water11. The previous ingredients were mixed to prepare the rearing media. By reaching pupation, around 100 pupae from each location were transferred into a cage (40×40×40 cm) provided with wire net from three sides to allow gas exchange as well as keeping adults inside the cage, while it provided with a hole covered with piece of cloth from the fourth side to allow dealing with insect. After adult emergence, they fed on powdered milk mixed with sugar (1:1) and cotton wick soaked with water was provided in a separate plastic cup. All the insects were maintained at 27±2°C, 60-65% RH and 14:10 h (light: dark) photoperiod.
Chemicals: Commercial grade formulation of lambda-cyhalothrin (Cycon 10% EC, Shoura Company, Egypt) was used for bioassays. α-Naphthyl acetate (α-NA), β-Naphthyl acetate (β-NA), diazoblue, sodium lauryl sulphate, α-naphthol, β-naphthol, methyl n butyrate (MeB), alkaline hydroxylamine, NaOH, HCl, FeCl3, 1-chloro 2,4-dinitrobenzene (CDNB), ethanol, P-nitroanisole o-demethylation, NADPH, glucose-6-phosphate, glucose-6-phosphate dehydrogenase (G-6-P-D), acetone, CHCl3, ethanol, bovine serum albumin (BSA) were products of sigma chemical Co (St Lois, MO, USA). Bovine albumin standard was purchased from Stanbio laboratory (Texas, USA). Coomassie brilliant blue G-250 was from sigma (sigma chemical Co.). P-nitroanisole (purity 97%) was obtained from Ubichem Ltd. (Hampshire), while nicotinamide adenine dinucleotide phosphate (reduced form, NADPH) was from BDH chemicals Ltd. (Poole, England).
Bioassay: Bioassays were performed in the Bioassay laboratory at Plant Protection Department, Zagazig University during 2017 and 2018 according to Kristensen and Jespersen12 with some modifications. Serial dilutions of 6 concentrations ranging from 50-350 μg mL–1 were prepared from lambda-cyhalothrin in distilled water (dH2O).
The larval rearing medium (as mentioned previously) was mixed with each insecticide concentration in 3 replicates for each concentration. Fifty 3rd instar larvae were used per each replicate with total number of 150 larvae for each concentration. Mortality was recorded after 72 h from treatment at conditions of 27±2°C, 60-65% RH and 14:10 h (light:dark) photoperiod. All larvae that were unable to develop into pupae were considered died.
Biochemical determination
Larval homogenate for protein and enzyme determination: Larval homogenates were prepared as described by Zhang et al.13. Larvae were homogenized in distilled water (50 mg mL–1). Larval homogenates were centrifuged at 8000 rpm for 15 min at 4°C. The supernatants, which were referred as enzymes extract, were stored at -20°C until used for enzyme activity determination within 2 weeks of extraction.
Total protein assay: Total proteins were determined by the method of Bradford14. Protein reagent was prepared by dissolving 100 mg of Coomassie Brilliant blue G-250 in 50 mL 95% ethanol. To this solution 100 mL 85 % (w/v) phosphoric acid was added. The resulting solution was diluted to a final volume of 1 L. Sample solution (50 μL) or 50 μL of serial concentrations containing 10-100 μg bovine serum albumin for preparation of standard curve were pipetted into test tubes. The volume in the test tube was adjusted to 1 mL with phosphate buffer (0.1 M, pH 6.6). Five milliliters of protein reagent were added to test tube and the contents were vortexed. The absorbance at 595 nm using Double beam ultraviolet/visible spectrophotometer (spectronic 1201, Milton Roy Co., USA) was measured after 2 min and before 1 h against blank. The blank was prepared by mixing 1 mL of phosphate buffer with 5 mL protein reagent.
General esterase assay: General esterase activity was determined according to Van Asparen15 using α-NA or β-NA as substrates. The reaction mixture consisted of 5 mL substrate solution (3×10–4 M α-NA or β-NA, 1% acetone and 0.1 M phosphate buffer, PH 7) and 20 μL of larval homogenate. The mixture was incubated for 15 min at 27°C and then 1 mL of diazoblue color reagent (prepared by mixing 2 parts of 1% diazoblue B and 5 parts of 5% sodium lauryl sulphate) was added. The developed color was measured at 600 or 555 nm. Double beam ultraviolet/visible spectrophotometer (spectronic 1201, Milton Roy Co., USA) was used for α-and β-naphthol produced from hydrolysis of the substrate, respectively. The α-and β-naphthol standard curves were prepared by dissolving 20 mg α-or β-naphthol in 100 mL phosphate buffer, PH 7 stock solution. Ten milliliters of stock solution were diluted up to 100 mL using phosphate buffer. Aliquots of 0.1, 0.2, 0.4, 0.8 and 1.6 mL of diluted solution (equal to 2, 4, 8, 16 and 32 μg naphthol) were pipetted into test tubes and completed to 5 mL by phosphate buffer. One milliliter of diazoblue reagent was added and the developed color was measured as mentioned before.
Carboxylesterase assay: Carboxylesterase activity was measured according to the method described by Simpson et al.16 using methyl n-butyrate (MeB) as a substrate. The reaction mixture contained 200 μL enzyme solution, 0.5 mL 0.067 M phosphate buffer, pH 7 and 0.5 mL MeB (4 mM). The test tubes were incubated at 37°C for 30 min. One milliliter of alkaline hydroxylamine (2 M hydroxylamine chloride and 3.5 M NaOH) was added to the test tubes and then 0.5 mL of HCl (1 part of concentration HCl and 2 parts of dH2O) was added. The mixture was shaken vigorously and allowed to stand for 2 min. Half milliliter of ferric chloride solution (0.9 M FeCl3 in 0.1 M HCl) was added and mixed well. The decrease in MeB resulting from hydrolysis by carboxylesterase was read at 515 nm, using Double beam ultraviolet/visible spectrophotometer (spectronic 1201, Milton Roy Co., USA).
Mixed function oxidase (MFO) assay: P-nitroanisole O-demethylation was assayed to determine the mixed function oxidase activity according to the method of Hansen and Hodgson17 with slight modification. The standard incubation mixture contained 1 mL sodium phosphate buffer (0.1 M, pH 7.6), 1.5 mL enzyme solution, 0.2 mL NADPH, (Final concentration 1 mM), 0.2 mL glucose-6-phosphate (G-6-P, final concentration, 1 mM) and 50 μg glucose-6-phosphate dehydrogenase (G-6-P-D). Reaction was initiated by the addition of P-nitroanisole in 10 μL of acetone to give a final concentration of 0.8 mM and incubated for 30 min at 37°C. Incubation period was terminated by addition of 1 mL HCl (1N). P-nitrophenol was extracted with CHCl3 and 0.5 N NaOH and absorbance of NaOH solution was measured at 405 nm using Double beam ultraviolet/visible spectrophotometer (spectronic 1201, Milton Roy Co., USA). An extinction coefficient of 14.28 mM cm–1 was used to calculate 4-nitrophenol concentration.
Glutathione S-transferase assay: Glutathione S-transferase (GST) catalyzes the conjugation of reduced glutathione (GSH) with 1-chloro 2,4-dinitrobenzene (CDNB) via the-SH group of glutathione. The conjugate, S-(2,4-dinitro-phenyl)-L-glutathione could be detected as described by the method of Habig et al.18. The reaction mixture consisted of 1 mL of the potassium salt of phosphate buffer (PH 6.5), 100 μL of GSH and 200 μL of larval homogenate. The reaction started by the addition of 25 μL of the substrate CDNB solution. The concentration of both GSH and CDNB was adjusted to be 5 mM and 1 mM, respectively. Enzyme and reagents were incubated at 30°C for 5 min. The increment in absorbance at 340 nm using Double beam ultraviolet/visible spectrophotometer (spectronic 1201, Milton Roy Co., USA) was recorded against blank containing all reagents except the enzyme solution to determine the nanomole substrate conjugated/min/larva using a molar extinction coefficient of 9.6 mM cm–1.
Statistical analysis: To estimate parameters of insect mortality in bioassay, the probit analysis program19 was used which corrected against control mortality20. Data of biochemical parameters were analyzed using SPSS program 14 for windows. Least significant difference (LSD) was used through ANOVA at p<0.05.
RESULTS AND DISCUSSION
Bioassay experiments: Results of M. domestica susceptibility are presented in Table 1. In comparison with laboratory strain, populations collected from Kafr Sakr showed the highest resistance to lambda-cyhalothrin (10.6 folds) at LC50. While at LC90, there are no high differences among populations collected as they showed from 2.3 fold (San El-Hagar population) to 3.1 fold of resistance (Abo Kabir population) comparing to laboratory strain. Regression line slope showed high slope value in Kafr Sakr population (5.32) which indicating homogeneity among individuals and low slope value in Zagazig population (2.82) comparing with (1.47) for laboratory strain.
Biochemical assays: Results of total protein concentration (Fig. 1) showed that populations collected from Abo Kabir and Zagazig recorded the highest significant concentration followed by kafr sakr, while San El-Hagar and Abo Hammad populations had the lowest protein concentration which was not significantly different with laboratory strain.
Results of esterase activity (Table 2) showed that populations of San El-Hagar and Kafr Sakr are significantly higher in esterase activity with 1.4 fold in comparison with laboratory strain when α-NA was used as a substrate. While the lowest activity of esterase was recorded in the population collected from Zagazig when α-NA was used as a substrate. When β-NA was used as a substrate, the same trend in esterase activity was noted except in Zagazig population which showed non-significant activity with San El-Hagar and significant activity with the rest populations and laboratory strain.
The highest significant activity of carboxylesterase (Table 3) was recorded in Zagazig population which showed 2.58-times higher than what recorded in laboratory strain. Kafr Sakr population showed 1.18-time higher in carboxylesterase activity comparing with laboratory strain followed by San El-Hagar which was non-significantly different with Abo Hammad, Abo Kabir and laboratory strain.
Results of MFO activity (Fig. 2) showed markedly higher in the activity of enzyme in both Zagazig and Abo Hammad populations. The lowest activity of MFO was recorded in both San El-Hagar population and laboratory strain. Activity in Abo Kabir and Kafr Sakr followed the activity recorded in Zagazig and Abo Hammad populations.
| Table 1: | Susceptibility of Musca domestica larvae of different localities in Sharkia Governorate to lambda-cyhalothrin |
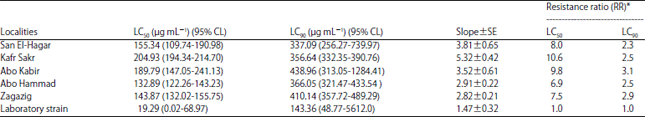 | |
Number of larvae used for each experiment/location = 50, experiments were repeated 3 times, *Values of resistance ratio were calculated by dividing LC50 or LC90 of the respective population by LC50 or LC90 of the laboratory strain | |
| Table 2: | Esterase activity in Musca domestica larvae of different locations from Sharkia Governorate |
 | |
*Values are shown as means±SD of 3 replicates, numbers followed by different letters within the same column indicate that data are statistically different at p<0.05, | |
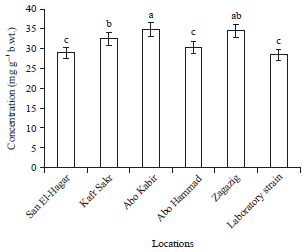 | |
| Fig. 1: | Total protein concentration in Musca domestica larvae of different locations from Sharkia Governorate |
Different letters on bars indicate that there are significant differences at p<0.05 | |
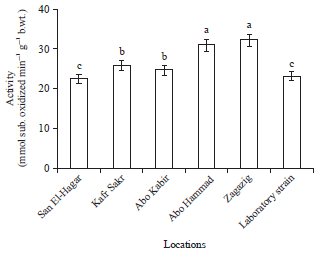 | |
| Fig. 2: | Mixed function oxidase activity in Musca domestica larvae of different locations from Sharkia Governorate |
Different letters on bars indicate that there are significant differences at p<0.05 | |
| Table 3: | Carboxylesterase activity in Musca domestica larvae of different locations from Sharkia Governorate |
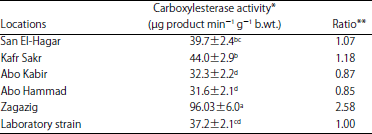 | |
*Values are shown as Means±SD of 3 replicates, the activity of carboxylesterase was determined as μg product (enzyme substrate) min–1 g–1 b.wt., numbers followed by different letters within the same column indicate that data are statistically different at p<0.05, **Ratio is calculated as mean values of collected populations/mean value of laboratory strain | |
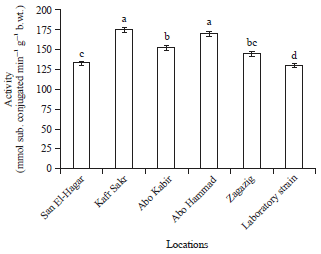 | |
| Fig. 3: | Glutathione S-transferase activity in Musca domestica larvae of different locations from Sharkia Governorate |
Different letters on bars indicate that there are significant differences at p<0.05 | |
Results of glutathione S-transferase activity (Fig. 3) showed that the highest significant activity was recorded in both kafr sakr and Abo Hammad populations, while the lowest significant activity was recorded in the laboratory strain which showed non-significant differences with Zagazig and San El-Hagar populations. The activity of GST in Abo Kabir population followed the activity in Kafr sakr and Abo Hammad populations.
Musca domistica susceptibility: Monitoring house fly susceptibility is important for the management control methods of this pest without harming the environment. Bioassay results showed 10.6 fold of tolerance in Kafr Sakr population to lambda-cyhalothrin. This result is similar to what Khan and Akram21 found when they tested the susceptibility of a field strain collected from Pakistan against different insecticides. Their bioassay experiments resulted from 5.9-9.7 fold of resistance of newly collected field strain of house fly toward pyriproxyfen and diflubenzuron, respectively. Also in support of the current results, different levels of resistance were reported in the freshly collected field strain of house fly which showed from low resistance (2 fold (to very high levels of resistance)>1219 fold (depending on insecticides tested13,22-27 ensuring carful use and regular monitoring of insect susceptibility in the environment toward different applied insecticides.
Activity of metabolic enzymes: Insects are naturally develop resistance against insecticides due to excessive use which promotes the development of high levels of resistance. Current results showed that different metabolic enzymes depending on the site of collection were higher in activity comparing with the reference strain with emphasis on carboxylesterase and MFO. In agreement with these results, different studies have demonstrated that metabolic mechanisms (Esterases, MFOs and GSTs) are responsible for pyrethroid resistance due to their major role in detoxification of insecticides in a number of insect pests28-30. The increased in enzymes activity reported in the current study including esterase and/or cytochrome P450 monooxygenase reflect the major role of these enzymes as a metabolic mechanism to pyrethroid resistance in insect pests including house fly as previously implicated31-33. The high in activity of the determined enzymes according to the site of population might be due to exposure to other insecticides that metabolized by the same enzymes leading to cross resistance against different other insecticides as what previously reported with fenvalerate, cyhalothrin, cypermethrin, acephate, chlorpyrifos and methomyl34-36.
On the other hand, a number of studies stated that cytochrome P450, GST and esterase enzymes not to be responsible for the main resistance mechanisms and other resistance mechanisms might be involved22,24-25,37 which vary depending on different factors such as the insect species, the degree of pesticide resistance, spatial and temporal variations, or combination of these factors37. Taking into consideration insect species, repeated use of insecticides and the nature of habitats, enzymes determined in the current study might be good indicators for showing resistance development of house fly toward a specific insecticide.
CONCLUSION
It is important to screen the susceptibility of M. domestica to insecticides use from time to time in order to develop strategies for resistance management. In this manner the susceptibility of M. domistica from locations in Sharkia Governorate was determined against the most frequent insecticide use. It was noted from bioassay experiments that house fly was slight to moderate resistant to lambda-cyhalothrin (2.3-10.6 fold). The levels of resistance was associated with ~3 times higher in carboxylesterase activity comparing with <1.4-times in MFO and GST activity indicating the vital role of esterase enzymes in insect resistance.
SIGNIFICANCE STATEMENT
This study discovers the susceptibility status of house fly populations from Sharkia Governorate, Egypt to lambda-cyhalothrin and the possible role of carboxylesterase in resistance development that can be beneficial for applying the most suitable control strategies keeping in view the increasing problem of insecticide resistance and the effect on environmental pollution. This study will help the researcher to cover the critical areas of the possibility of repeating use of insecticides in house fly control. Thus a new theory on application of different control programs including those depend on the rotation of insecticides application may be arrived at.
REFERENCES
- Pavela, R., 2008. Insecticidal properties of several essential oils on house fly (Musca domestica L.). Phytother. Res., 22: 274-278.
PubMed - Khan, H.A.A., S.A. Shad and W. Akram, 2012. Effect of livestock manures on the fitness of house fly, Musca domestica L. (Diptera: Muscidae). Parasitol. Res., 111: 1165-1171.
CrossRefDirect Link - Zhu, F., L. Lavine, S. O’Neal, M. Lavine, C. Foss and D. Walsh, 2016. Insecticide resistance and management strategies in urban ecosystems. Insects, Vol. 7.
CrossRefDirect Link - Shafer, T.J., D.A. Meyer and K.M. Crofton, 2005. Developmental neurotoxicity of pyrethroid insecticides: Critical review and future research needs. Environ. Health Perspect., 113: 123-136.
CrossRefPubMedDirect Link - Abbas, N., H.A.A. Khan and S.A. Shad, 2014. Resistance of the house fly Musca domestica (Diptera: Muscidae) to lambda-cyhalothrin: Mode of inheritance, realized heritability and cross-resistance to other insecticides. Ecotoxicology, 23: 791-801.
CrossRefDirect Link - Kaufman, P.E., S.C. Nunez, R.S. Mann, C.J. Geden and M.E. Scharf, 2010. Nicotinoid and pyrethroid insecticide resistance in houseflies (Diptera: Muscidae) collected from Florida dairies. Pest Manage. Sci., 66: 290-294.
CrossRefDirect Link - Bass, C. and M.F. Linda, 2011. Gene amplification and insecticide resistance. Pest Manage. Sci., 67: 886-890.
CrossRefDirect Link - Aïzoun, N., R. Aïkpon, G.G. Padonou, O. Oussou and F. Oké-Agbo et al., 2013. Mixed-function oxidases and esterases associated with permethrin, deltamethrin and bendiocarb resistance in Anopheles gambiae sl in the South-North transect Benin, West Africa. Parasit. Vect., Vol. 6.
CrossRefDirect Link - Tian, L., C. Cao, L. He, M. Li and L. Zhang et al., 2011. Autosomal interactions and mechanisms of pyrethroid resistance in house flies, Musca domestica. Int. J. Biol. Sci., 7: 902-911.
Direct Link - Khan, H.A.A., W. Akram and S.A. Shad, 2013. Resistance to conventional insecticides in Pakistani populations of Musca domestica L.(Diptera: Muscidae): A potential ectoparasite of dairy animals. Ecotoxicology, 22: 522-527.
CrossRefDirect Link - Khan, H.A.A., W. Akram, S.A. Shad, M. Razaq, U. Naeem-Ullah and K. Zia, 2013. A cross sectional survey of knowledge, attitude and practices related to house flies among dairy farmers in Punjab, Pakistan. J. Ethnobiol. Ethnomed., Vol. 9.
CrossRefDirect Link - Kristensen, M. and J.B. Jespersen, 2003. Larvicide resistance in Musca domestica (Diptera: Muscidae) populations in Denmark and establishment of resistant laboratory strains. J. Econ. Entomol., 96: 1300-1306.
CrossRefDirect Link - Zhang, L., X. Gao and P. Liang, 2007. Beta-cypermethrin resistance associated with high carboxylesterase activities in a strain of house fly, Musca domestica (Diptera: Muscidae). Pestic. Biochem. Physiol., 89: 65-72.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Hansen, L.G. and E. Hodgson, 1971. Biochemical characteristics of insect microsomes: N and O-demethylation. Biochem. Pharmacol., 20: 1569-1578.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Khan, H.A.A. and W. Akram, 2017. Cyromazine resistance in a field strain of house flies, Musca domestica L.: Resistance risk assessment and bio-chemical mechanism. Chemosphere, 167: 308-313.
CrossRefDirect Link - Li, J., Q. Wang, L. Zhang and X. Gao, 2012. Characterization of imidacloprid resistance in the housefly Musca domestica (Diptera: Muscidae). Pestic. Biochem. Physiol., 102: 109-114.
CrossRefDirect Link - Khan, H.A.A., W. Akram, J. Iqbal and U. Naeem-Ullah, 2015. Thiamethoxam resistance in the house fly, Musca domestica L.: Current status, resistance selection, cross-resistance potential and possible biochemical mechanisms. Plos One, Vol. 10.
CrossRefDirect Link - Khan, H.A.A., W. Akram, T. Khan, M.S. Haider, N. Iqbal and M. Zubair, 2016. Risk assessment, cross-resistance potential and biochemical mechanism of resistance to emamectin benzoate in a field strain of house fly (Musca domestica Linnaeus). Chemosphere, 151: 133-137.
CrossRefDirect Link - Li, Q., J. Huang and J. Yuan, 2018. Status and preliminary mechanism of resistance to insecticides in a field strain of housefly (Musca domestica, L). Rev. Brasil. Entomol., 62: 311-314.
Direct Link - Cetin, H., O. Kocak, E. Oz, S. Koc, Y. Polat and K. Arıkan, 2019. Evaluation of some synthetic pyrethroids and piperonyl butoxide combinations against Turkish house fly (Musca domestica L.) populations. Pak. J. Zool., 51: 703-707.
Direct Link - Wang, J.N., J. Hou, Y.Y. Wu, S. Guo, Q.M. Liu, T.Q. Li and Z.Y. Gong, 2019. Resistance of house fly, Musca domestica L. (Diptera: Muscidae), to five insecticides in Zhejiang province, China: The situation in 2017. Canad. J. Infect. Dis. Med. Microbiol., Vol. 2019.
CrossRefDirect Link - Liu, X., P. Liang, X. Gao and X. Shi, 2006. Induction of the cytochrome P450 activity by plant allelochemicals in the cotton bollworm, Helicoverpa armigera (Hübner). Pestic. Biochem. Physiol., 84: 127-134.
CrossRefDirect Link - Lai, T., J. Li and J. Su, 2011. Monitoring of beet armyworm Spodoptera exigua (Lepidoptera: Noctuidae) resistance to chlorantraniliprole in China. Pestic. Biochem. Physiol., 101: 198-205.
CrossRefDirect Link - Gao, C.F., S.Z. Ma, C.H. Shan and S.F. Wu, 2014. Thiamethoxam resistance selected in the western flower thrips Frankliniella occidentalis (Thysanoptera: Thripidae): Cross-resistance patterns, possible biochemical mechanisms and fitness costs analysis. Pestic. Biochem. Physiol., 114: 90-96.
CrossRefDirect Link - Jones, C.M., K.A. Haji, B.O. Khatib, J. Bagi and J. Mcha et al., 2013. The dynamics of pyrethroid resistance in Anopheles arabiensis from Zanzibar and an assessment of the underlying genetic basis. Parasit. Vect., Vol. 6.
CrossRefDirect Link - Demkovich, M., C.E. Dana, J.P. Siegel and M.R. Berenbaum, 2015. Effect of piperonyl butoxide on the toxicity of four classes of insecticides to navel orangeworm (Amyelois transitella)(Lepidoptera: Pyralidae). J. Econ. Entomol., 108: 2753-2760.
CrossRefDirect Link - Safi, N.H.Z., A.A. Ahmadi, S. Nahzat, S.P. Ziapour and S.H. Nikookar et al., 2017. Evidence of metabolic mechanisms playing a role in multiple insecticides resistance in Anopheles stephensi populations from Afghanistan. Malaria J., Vol. 16.
CrossRefDirect Link - Wan, P.J., X.Q. Shi, Y. Kong, L.T. Zhou, W.C. Guo, T. Ahmat and G.Q. Li, 2013. Identification of cytochrome P450 monooxygenase genes and their expression profiles in cyhalothrin-treated Colorado potato beetle, Leptinotarsa decemlineata. Pestic. Biochem. Physiol., 107: 360-368.
CrossRefDirect Link - Rasool, A., N. Joußen, S. Lorenz, R. Ellinger and B. Schneider et al., 2014. An independent occurrence of the chimeric P450 enzyme CYP337B3 of Helicoverpa armigera confers cypermethrin resistance in Pakistan. Insect Biochem. Mol. Biol., 53: 54-65.
CrossRefDirect Link - Rodrigues, A.R., H.A. Siqueira and J.B. Torres, 2014. Enzymes mediating resistance to lambda-cyhalothrin in Eriopis connexa (Coleoptera: Coccinellidae). Pestic. Biochem. Physiol., 110: 36-43.
CrossRefDirect Link - Gong, Y.J., Z.H. Wang, B.C. Shi, Z.J. Kang and L. Zhu et al., 2013. Correlation between pesticide resistance and enzyme activity in the diamondback moth, Plutella xylostella. J. Insect Sci., Vol. 13.
CrossRefDirect Link