
Ihsan Nurkomar
Laboratory of Biological Control, Department of Plant Protection, Faculty of Agriculture, Bogor Agricultural University, Jalan Kamper Kampus, IPB Dramaga, 16680 Bogor, Indonesia
Nurul Novianti
Laboratory of Biological Control, Department of Plant Protection, Faculty of Agriculture, Bogor Agricultural University, Jalan Kamper Kampus, IPB Dramaga, 16680 Bogor, Indonesia
Pudjianto
Department of Plant Protection, Faculty of Agriculture, Bogor Agricultural University, Jalan Kamper Kampus, IPB Dramaga, 16680 Bogor, Indonesia
Syafrida Manuwoto
Department of Plant Protection, Faculty of Agriculture, Bogor Agricultural University, Jalan Kamper Kampus, IPB Dramaga, 16680 Bogor, Indonesia
Damayanti Buchori
Department of Plant Protection, Faculty of Agriculture, Bogor Agricultural University, Jalan Kamper Kampus, IPB Dramaga, 16680 Bogor, Indonesia
LiveDNA: 62.16628
Journal of Entomology
Year: 2017 | Volume: 14 | Issue: 3 | Page No.: 128-135







ABSTRACT
Background and Objective: Diaphania indica (D. Indica) is a major pest of certain cucurbit plants. The species has been classified as minor pest in Indonesia, but has the potential to become a major pest. Apanteles taragamae, (A. taragamae) a larval parasitoid of D. indica, holds promise as a biological control agent of this cucumber pest. The objective of this study was to understand the effectiveness of A. taragamae as biological control agent for D. indica. Methodology: The effectiveness of A. taragamae was studied through its life cycle, egg maturation, fitness characteristics and functional responses. The biological data were analyzed by one-way analysis of variance (ANOVA) followed by Tukey’s test. The relationship between body size and fecundity as well as functional responses was analyzed by simple regression analysis. Results: The incubation period of egg was 1-2 days. There are 3 instars stages of larva, the duration of 1st, 2nd and 3rd instar are 2, 3 and 2 days, respectively. Total larval phase is about 7 days, pupae formed in a few h after pre-pupa period. Pupae period is about 7 days. After emergence, female already in possession of a supply of mature eggs inside the ovaries which was significantly different for females of different ages (p<0.0001). Female possess high potential fecundity which highly correlates to her body size. In addition, A. taragamae shows host density-dependent responses to its host D. indica. Conclusion: The results indicate that A. taragamae is an effective koinobiont parasitoid species and biologically well-adapted to its host D. indica.
PDF Abstract XML References Citation
Received: January 05, 2017;
Accepted: March 16, 2017;
Published: April 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ihsan Nurkomar, Nurul Novianti, Pudjianto, Syafrida Manuwoto and Damayanti Buchori, 2017. Biology and Fitness Characteristics of Apanteles taragamae Viereck (Hymenoptera: Braconidae). Journal of Entomology, 14: 128-135.
DOI: 10.3923/je.2017.128.135
URL: https://scialert.net/abstract/?doi=je.2017.128.135
DOI: 10.3923/je.2017.128.135
URL: https://scialert.net/abstract/?doi=je.2017.128.135
INTRODUCTION
The role of parasitoids in regulating host populations depends on their compatibility with the biological characteristics of their selected host and their overall fitness as a parasitoid. In general, fitness is a term used by biologists to describe how well an individual performs in a population, in terms of maximal production of healthy offspring1. Several studies have described how parasitoid fitness relates to biological traits2. For instance, maternal egg size of Lysiphlebus fabarum (Hymenoptera: Braconidae) is positively correlated to her progeny developmental time and egg load3. Adult body size of Diaeretiella rapae (Hymenoptera: Aphidiidae) is positively correlated with the number of offspring and the progeny sex ratio4.
Diaphania indica is an occasionally serious pest of certain Cucurbitaceae species5 and is also widespread in Japan, Korea, China, Taiwan, Tropical Asia, Africa, the Netherlands, some Pacific islands and the United States6. Infestation at rates of one D. indica larva/leaf in cucumber plants can cause 10% yield loss7. However, the species has so far been classified as a minor pest. It has a rapid development rate, high survival rate and a large reproductive capacity8. Thus, this species could potentially be a major pest and cause significant large-scale crop damage. A. taragamae has been identified as a promising candidate for biological control of D. indica9,10. However, known information about this parasitoid is still limited.
To address potential threat of D. indica, the key aspects of parasitoid biology and ecology should be understood or evaluated to ensure the success of any biological control program11, since D. indica’s use of cucurbit leaves to create closed shelters by folding leaves around their bodies represents a primary limiting factor for effective chemical pest control8. The biological traits of A. taragamae were reported previously by Peter and David10. However, the relationship between the biology of A. taragamae and its fitness characteristics remain unknown. The present research aims to studied the effectiveness of A. taragamae as biological control agent for D. indica through study the aspects of biology and fitness characteristics of A. taragamae, including life cycle and egg maturation, the relationship between adult body size and fecundity, as well as the aspect of functional responses.
MATERIALS AND METHODS
Insects: Larvae and pupae of both the target parasitoid and host species were collected from a cucumber field in Dramaga, Bogor, Indonesia, from March, 2015-January, 2016. D. indica samples were placed in a cylindrical plastic cage (10 cm in diameter, 30 cm in high) containing bottled-water and cucumber leaves for oviposition and cotton moistened with 20% honey solution as a food source. Any leaves holding egg masses were collected daily and transferred to another cylindrical plastic cage (15 cm in diameter, 10 cm high) until the first instar developed. These were then removed to plastic containers (25×15×5 cm) and reared with a natural diet, i.e., 5-10 cucumber leaves, until pupation. The pupae were transferred to Petri dish (86×13 mm) and kept there until emergence of adults.
Collected A. taragamae cocoon clusters were placed in a 50 mL test tube until emergence of adults. Honey droplets were provided as food. The collected insects of both species were kept in a rearing cabinet in laboratory conditions (25±1°C, 90±10% RH and L16:D8 photoperiod). All parasitoid adults used in this experiment were directly collected from the field, due to the difficulty of rearing successive generations of parasitoids in the lab.
Life cycle of A. taragamae: To study the life cycle of A. taragamae, an experiment was performed exposing ten 3-days-old first instar larvae of D. indica to one mated A. taragamae female in a Petri dish (86×13 mm). In the preliminary tests, initial parasitization of larvae and the formation of parasitoid cocoon clusters elapsed within 14 days. Therefore, 14 Petri dishes were prepared for a total of 14 tests, using a new parasitoid female for each test. The parasitoid females were added to their respective dishes at the same time, left for 24 h and then removed. Each day thereafter, 10 parasitized larvae were removed from one of the dishes, sequentially, to yield samples 1 days old, 2 days old, 3 days old and so on, through 14 days old. The parasitized larvae removed each day, were dissected to observe the developmental stage of the parasitoid inside the host’s body.
Longevity of A. taragamae adults was measured using individuals newly emerged in the lab from field collected colonies. A total of 50 males and 50 females were collected and placed in groups of 10 (segregated by sex) into a 50 mL test tube (N = 10 tubes). In total, 50 females and 50 males was used. Honey droplets were provided as a food source. Adult parasitoids were reared until death. The number and sex of adult parasitoids deceased each day was recorded and the average longevity of adults were calculated for each sex.
Egg maturation: To study egg maturation time in A. taragamae, adult female parasitoids of different ages: 0, 1, 2, 3, 4, 5, 6, 12, 24, 48, 72, 96, 120, 144, 168, 192, 216, 240 and 264 h after eclosion were dissected. The parasitoids were paralyzed by storing them at a cold temperature (±10°C) for 1 h prior to dissection. While still alive, the abdomen of each individual was dissected using a stemmed micro-needle under a stereo microscope. The number of mature eggs inside the parasitoid ovary was counted using a mechanical manual hand counter. Ten females in each age group were dissected, as replication.
Fitness characteristics of A. taragamae: The various fitness characteristics of A. taragamae, including parasitism rate, total number of oviposited eggs, daily egg oviposition, number of eggs remaining inside the ovary, longevity and sex ratio of progeny were studied. The experiment involved daily introduction of one 2-day-old mated female A. taragamae to ten 3-day-old host larvae. The larvae were changed daily but the same female parasitoid was introduced each day, until the female parasitoid died. The host larvae were kept in a cylindrical plastic cage (10 cm in diameter, 15 cm high) and given 2 cucumber leaves as food. For each introduction, the parasitoid was released into the cage and allowed to parasitize for 24 h. The cage, host larvae and food (cucumber leaves) were changed each day for the next exposure. Parasitized larvae were reared in a new cage and supplied with new cucumber leaves everyday until parasitoid cocoon clusters formed.
As parasitoid cocoons formed, each cocoon cluster was placed in a 50 mL test tube until emergence. Honey droplets were provided as food. After emergence, the progeny sex ratio was calculated. Occasionally, parasitized larvae died before a parasitoid cocoon cluster had formed. In this case, the larvae were dissected to determine the number of pre-imaginal stage parasitoids within the hosts’ bodies. The results were tallied as oviposited eggs and included in those data.
The fitness parameters mentioned above were measured as appropriate throughout the experiment. For example, daily oviposition and number of host larvae parasitized could be counted every day, while parasitoid longevity, total eggs oviposited and eggs remaining in the ovary could only be counted after the death of the female parasitoid. In addition to the fitness traits, physical measurements of the female parasitoids, including the length of wings, hind tibia and body was done, after the final day of exposure.
Functional responses of A. taragamae: To determine the effect of host density on the parasitism rate of D. indica by A. taragamae, a series of experiments were performed featuring different host densities. Groups of 10, 20, 30 and 40 three-days-old first instar D. indica larvae were each put in a Petri dish (86×13 mm) containing cucumber leaves. One 2-days-old mated female parasitoid was placed in each Petri dish along with the larvae for 24 h. Then the parasitoids were removed and the host larvae were dissected to determine the parasitism rate for each treatment tested. This process was repeated 10 times at each density level, using different female parasitoids, for a total of 40 replications and 40 mated female parasitoids used.
Statistical analysis: Data on the number of mature eggs inside the ovaries were analyzed by one-way ANOVA12 and calculated means were separated by using Tukey Honest Significant Difference test (Tukey HSD test, α = 5%) using SAS Statistic Analytical Software version 9.4. The relationship between body size and fecundity of A. taragamae was analyzed using simple linear regression analysis. As well as, the relationship between host density and the parasitism rate.
RESULTS
Life Cycle of A. taragamae: The life cycle of A. taragamae consists of the following life stages: Egg, larva, pre-pupa, pupa and adult (Table 1). Eggs develop for 1-2 days (Fig. 1a-c). Larvae pass through three instar stages i.e., first instar larvae which develop for 1-2 days (Fig. 1d) and the second (Fig. 1e) and the third instar larvae (Fig. 1f) which develop for 2-5 days and 2-3 days, respectively. A. taragamae remained the pre-pupae stage for only 5-8 h, before passing into the pupal stage lasting for 5-6 days until emergence. Female and male adult longevity averaged 17 and 18 days, respectively.
Egg maturation: The average number of mature eggs inside the ovaries was significantly different for females of different ages (F = 80.81, df = 18, p<0.0001) (Fig. 2).
| Table 1: | Duration of different stages in the life cycle of A. taragamae |
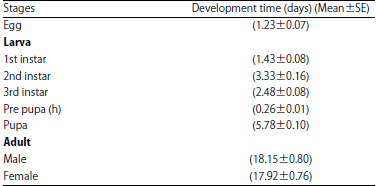 | |
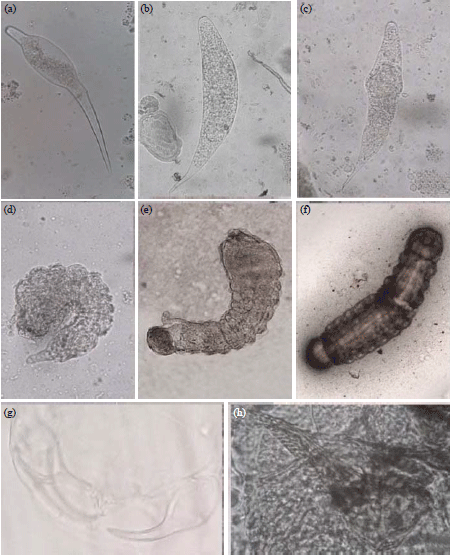 | |
| Fig. 1(a-h): | Developmental stages of A. taragamae, (a) Egg in the ovary, (b) Oviposited egg inside the host body, (c) Egg just before hatching, (d) First instar larva, (e) Second instar larva, (f) Third instar, (g) Mandible of first instar larva and (h) Mandible of third instar larva |
| Table 2: | Biological parameters of A. taragamae |
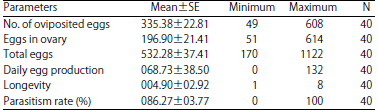 | |
The number increased in females aged 0-5 h after eclosion and then plateaued for ages 6-24 h after eclosion. The number of mature eggs inside the ovary increased again in females aged 48-72 h after eclosion and stabilized for those aged 96-144 h after eclosion. Thereafter, the average number of mature eggs decreased in females aged 168-264 h after eclosion.
Biological traits and fitness characteristics of A. taragamae: Adult A. taragamae females laid an average of 68.73 eggs distributed among 10 host larvae/day, for 4.90 days. This corresponds to a calculated average of 335.38 total oviposited eggs. On average, 196.90 eggs still remained inside the ovaries of the parasitoids after the test, bringing average total lifetime fecundity of studied females to 532.28 eggs (Table 2). The mature eggs harvested from this experiment were reared in the lab until adulthood and these adult progenies were examined to determine sex. The sex ratios of A. taragamae progeny calculated for the majority were male-biased.
Simple regression analysis showed a positive correlation between female body size (body length, p = 4.83e-07, wing length, p = 4.38e-06 and hind tibia length, p = 2.21e-07) and fecundity. There was also a positive correlation between female body size (body length, p = 6.71e-05, wing length, p = 3.05e-04 and hind tibia length, p = 3.18e-05) and number of eggs oviposited (Fig. 3).
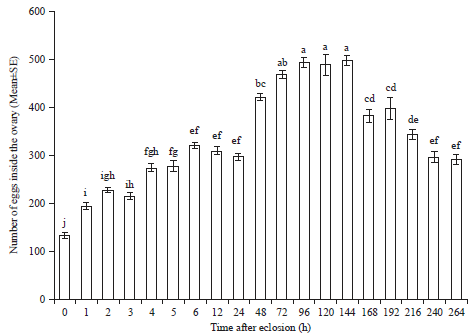 | |
| Fig. 2: | Effects of female age on the number of mature eggs inside the ovary. Each bar shows the mean number of mature eggs inside the ovary (Mean±S.E). Means followed by different letters are significantly different (Tukey HSD Test, α = 5%) |
Functional responses of A. taragamae: A. taragamae displayed host density-dependent functional responses within the range of host densities tested. The number of parasitized larvae increased linearly as host larval density increased (y = 8.37×-0.05, R2 = 0.9812, F = 741.4, P = 2e-16) (Fig. 4).
DISCUSSION
Observations of A. taragamae development show that the species follows holometabolan metamorphosis. Each pre-imaginal development stage of A. taragamae found inside the body of host larvae was distinguished, per methods used by Peter and David10. A. taragamae eggs collected were transparent and had a smooth surface (Fig. 1a-c). Each egg is acuminate and devoid of any sculpturing. The cephalic end is rounded and the caudal end slightly narrowed with a short curved pedicel. The eggs developed for 1-2 days, then hatched into larvae, a stage which consists of three instars. Each instar was distinguished by the number of abdominal segments and the shape of the mandible.
As described by Peter and David10, the first instar larvae floated freely out from the egg into a hemosol solution (Fig. 1d). These first instar larvae had 8 abdominal segments and a sickle-shaped mandible easily visible at the time of hatching (Fig. 1g). The appearance of the larva changes toward the end of this instar stage. Specifically, a cap-like swelling at the posterior end later invaginates to form the anal vesicle. The second instar larvae were more distinctly segmented (Fig. 1e). The body was vesiculate and the anal vesicle was fully developed and bladder-like. The head was not sclerotized and the mouth parts were not well developed. The mandibles were composed of fleshy lobes and were difficult to detect. The third instar larvae had 13 abdominal segments which were clearly visible (Fig. 1f). The anal vesicle was reduced considerably and had a shriveled appearance. The third instar larvae were white and the head was well sclerotized. The mouthparts were not visible externally. The mandibles are saw-like and prominent on the inner edge of the blad and invisible externally (Fig. 1h).
In normal larval development, the 3rd instar larvae will eclose out of the host body and spin a cocoon cluster for 5-8 h. The cocoons were bright-white in color, very compact, cylindrical and rounded at both ends. The cocoons developed for 5-6 days before the pupae eclosed into the adult stage. Emergent adults were small (2-4 mm in length), with a black body and transparent wings. The sex of adult individuals could be distinguished by body size, presence/absence of the ovipositor and length of antenna. Females were larger than the males, possessed an ovipositor and had a shorter antenna length. These also consistent with the previous study reported by Peter and David10.
The number of mature eggs inside the ovaries differs according to the age of the individual. Mature eggs can be found inside newly-emerged females just after eclosion. This suggests that A. taragamae is a pro-synovigenic parasitoid: A parasitoid that emerges into adulthood already in possession of a supply of mature eggs and which continues to produce additional mature eggs if/when the original egg supply is depleted13.
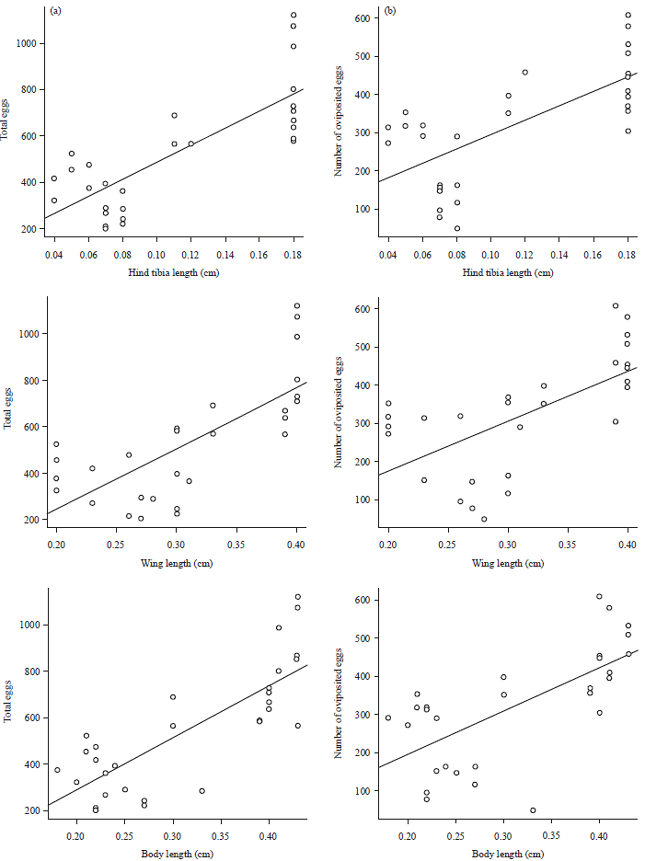 | |
| Fig. 3(a-b): | Relationship between body size parameters and (a) The total number of eggs produced and (b) The number of eggs oviposited, of A. taragamae. N = 29 |
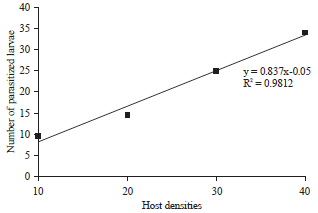 | |
| Fig. 4: | Relationship between larval density of host D. indica and number of host larvae parasitized by A. taragamae |
The number of mature eggs fluctuated among the females tested. This is because during the egg maturation experiment, females were not exposed to host larvae. Hegazi et al.14 said that host deprivation may result in a decrease over time in the number of eggs found in the ovaries after eclosion. Host deprivation may cause the depletion of eggs in the oviduct which is usually attributed to reabsorption of the eggs, known as Oosorption12.
Related to those results, female A. taragamae possess high potential fecundity. Potential fecundity is measured as the maximum egg production of a female parasitoid during her entire life. This potential fecundity includes the number of oviposited eggs and the number of eggs remaining inside the ovary at death. Variation in egg production can be influenced by availability and type of food and by the number of hosts available. Although the number of D. indica larvae exposed to A. taragamae was controlled (i.e., ten 3-days-old larvae) individual larvae may vary in quality. The female parasitoid will lay more eggs if the available host provides sufficient nutrition for egg development. Lebreton et al.15 explained that the number of eggs oviposited on each host varies, depending on the quality of the host. Similarly, Segoli et al.16 also confirmed that the quality of the host influences the decision of female to lay her eggs.
Female body size (length of thorax+abdomen), as well as the length of the forewings and hind tibia, were positively correlated to female fecundity for A. taragamae. Therefore, those physical parameters could serve as a useful indicator of the species fitness. Waschke et al.17 support this view that body size is associated with fitness in the field, it contributes directly to parasitoid ability to find hosts.
The parasitoid progeny sex ratio reared from laboratory colonies were skewed male. The preponderance of male progeny might be due to progeny derived from unfertilized females. Parasitoids may utilize a form of parthenogenetic reproduction which results in male offspring only. This is common in haplodiploidi hymenoptera18 such as in Cotesia glomerata19 and Gonatocerus ashmedi20.
CONCLUSION
A. taragamae is an effective koinobiont parasitoid and biologically well-adapted to its host D. indica. Females were highly fecund, a primary indicator of fitness and highly correlated with her body size. A. taragamae showed clear host-density dependence, such that parasitism rates were higher with higher host density.
SIGNIFICANCE STATEMENT
This study revealed the potential of using A. taragamae as a control option for the cucumber moth D. indica. This study will help the researchers to expose the ecological relationship between D. indica and A. taragamae its cryptic nature that contributes to their complexity in management.
ACKNOWLEDGMENTS
This study was supported by the Ministry of Research, Technology and Higher Education of Indonesia PMDSU Program No. 081/SP2H/PL/DIT.LITABMAS/VI/2015.
REFERENCES
- Roitberg, B.D., G. Boivin and L.E.M. Vet, 2012. Fitness, parasitoids and biological control: An opinion. Can. Entomol., 133: 429-438.
CrossRefDirect Link - Sarfraz, M., L.M. Dosdall and B.A. Keddie, 2009. Fitness of the parasitoid Diadegma insulare is affected by its host's food plants. Basic Applied Ecol., 10: 563-572.
CrossRefDirect Link - Ameri, M., A. Rasekh and J.P. Michaud, 2014. Body size affects host defensive behavior and progeny fitness in a parasitoid wasp, Lysiphlebus fabarum. Entomol. Exp. Applic., 150: 259-268.
CrossRefDirect Link - Kant, R., M.A. Minor, S.A. Trewick and W.R.M. Sandanayaka, 2012. Body size and fitness relation in male and female Diaeretiella rapae. BioControl, 57: 759-766.
CrossRefDirect Link - Pandey, P.N., 1975. Infestation of Diaphania indica Saund. on cucurbits. J. Applied Entomol., 79: 160-163.
CrossRefDirect Link - Peter, C. and B.V. David, 1991. Population dynamics of the pumpkin caterpillar, Diaphania indica (Saunders)(Lepidoptera: Pyralidae). J. Trop. Pest Manage., 37: 75-79.
CrossRefDirect Link - Schreiner, I.H., 1991. Damage threshold for Diaphania indica Saunders (Lepidoptera: Pyralidae) on cucumbers in Guam. J. Trop. Pest Manage., 37: 17-20.
CrossRefDirect Link - Peter, C. and B.V. David, 1991. Natural enemies of the pumpkin caterpillar Diaphania indica [Lepidoptera: Pyralidae] in Tamil Nadu. Entomophaga, 36: 391-394.
CrossRefDirect Link - Peter, C. and B.V. David, 1992. Biology of Apanteles taragamae Viereck (hymenoptera: Braconidae) a parasitoid of Diaphania indica (Saunders) (Lepidoptera: Pyralidae). Int. J. Trop. Insect Sci., 13: 7-17.
CrossRefDirect Link - Lupi, D., R. Favaro, C. Jucker, C.O. Azevedo, I.C.W. Hardy and M. Faccoli, 2017. Reproductive biology of Sclerodermus brevicornis, a European parasitoid developing on three species of invasive longhorn beetles. Biol. Control, 105: 40-48.
CrossRefDirect Link - Irvin, N.A. and M.S. Hoddle, 2009. Egg maturation, oosorption and wing wear in Gonatocerus ashmeadi (Hymenoptera: Mymaridae), an egg parasitoid of the glassy-winged sharpshooter, Homalodisca vitripennis (Hemiptera: Cicadellidae). Biol. Control, 48: 125-132.
CrossRefDirect Link - Hegazi, E.M., G.M. Abd El-Aziz, A.Y. El-Shazly and W.E. Khafagi, 2007. Influence of host age and host deprivation on longevity, egg load dynamics and oviposition rate in Microplitis rufiventris. Insect Sci., 14: 485-495.
CrossRefDirect Link - Lebreton, S., M. Labarussias, C. Chevrier and E. Darrouzet, 2009. Discrimination of the age of conspecific eggs by an ovipositing ectoparasitic wasp. Entomol. Exp. Applic., 130: 28-34.
CrossRefDirect Link - Segoli, M., T. Keasar, A. Bouskila and A.R. Harari, 2010. Host choice decisions in the polyembryonic wasp Copidosoma koehleri (Hymenoptera: Encyrtidae). Physiol. Entomol., 35: 40-45.
CrossRefDirect Link - De la Filia, A.G., S.A. Bain and L. Ross, 2015. Haplodiploidy and the reproductive ecology of Arthropods. Curr. Opin. Insect Sci., 9: 36-43.
CrossRefDirect Link - Zhou, Y., H. Gu and S. Dorn, 2006. Single-locus sex determination in the parasitoid wasp Cotesia glomerata (Hymenoptera: Braconidae). Heredity, 96: 487-492.
Direct Link - Lytle, J., A. Kapranas, D.J.W. Morgan, J.M. Nichols and J.G. Morse, 2012. Sex ratio dynamics of Gonatocerus ashmeadi, a parasitoid of the glassy-winged sharpshooter in California. Entomol. Exp. Applic., 145: 222-227.
CrossRefDirect Link