
A.A.A. El- Sayed
Plant Protection Research Institute, Agriculture Research Center, Dokki, Giza, Egypt
Journal of Entomology
Year: 2016 | Volume: 13 | Issue: 1-2 | Page No.: 26-32







ABSTRACT
The effect of LC20 of hexaflumuron on some biological aspects, egg maturation and ovary of the spiny bollworm, Earias insulana (Boisd.) were studied under controlled conditions of 26±1°C and 75±5 RH. The calculated LC20 and LC50 values of hexaflumuron were 17.42 and 72.75 ppm. The treatment newly hatched larvae with LC20 (17.42 ppm) elongated larval and pupal duration compared with control. Treatment significantly reduced mean numbers of deposited eggs and hatchability percentage compared with control. Hexaflumuron reduced number of immature eggs after emergence (20.56 eggs/ovariol) compared with 27.05 eggs/ovariol for untreated. No mature eggs were present in the female moths ovarioles after emergence of treated and untreated. Mature eggs were found after 36 h and 3 days in untreated and treated, respectively. The highest numbers of mature eggs 3.63 and 6.32 eggs/ovariol were recorded after four days from emergence in treated and untreated, respectively. Hexaflumuron reduced the ovarian weight and length, size of basal eggs and number of spermatophore/female compared with untreated. The LC20 of hexaflumuron reduced significantly larval content of total carbohydrate, lipid and soluble protein.
PDF Abstract XML References Citation
Received: October 29, 2015;
Accepted: December 17, 2015;
Published: February 15, 2016
How to cite this article
A.A.A. El- Sayed, 2016. Effect of Sublethal Dose of Hexaflumuron on the Ovary and Egg Maturation of Spiny Bollworm, Earias insulana (Boisd.). Journal of Entomology, 13: 26-32.
DOI: 10.3923/je.2016.26.32
URL: https://scialert.net/abstract/?doi=je.2016.26.32
DOI: 10.3923/je.2016.26.32
URL: https://scialert.net/abstract/?doi=je.2016.26.32
INTRODUCTION
The spiny bollworm, Earias insulana (Boisd.) is one of the most dangerous pests attacking many plants such as cotton, okra and maize. Hexaflumuron is a benzoylphenylurea derivative that interferes with chitin synthesis during molting. This mode of action effectively controls the immature stages of insects, which are the most damaging stages in many insect pests where the larval stage cause the maximum harm to crops (Retnakaran et al., 1985). Horowitz et al. (1992) found that LC50 of hexaflumuron was 0.25 ppm for newly hatched larvae of the spiny bollworm. Eggs dipping in concentration of 50 ppm resulted in suppression of 80% egg hatch. Perveen and Miyata (2000) found that treated the fifth instar larvae of Spodoptera litura topically with sublethal doses of chlorfluazuron (1.00 ng/larva or 3.75 ng/larva) reduced ovarian weight and number of mature eggs in adults compared with untreated. Also, disrupt growth and development of oocytes by significant affecting the size of basal oocytes and protein content of ovarian was significant reduced. Soltani and Soltani-Mazouni (1997) cleared that diflubenzuron topically application (0.5 ug/insect) to Cydia pomonella at pupal ecdysis disturbed growth and development of oocytes and reduced size of basal oocytes, the protein content per ovary and the number of oocytes per ovary in newly emerged adults. Khebbeb et al. (1997) found that diflubenzuron decreased the overall lipid level in the fat body in the adult females of Tenebrio molitor treated. In adult females, the major function of the fat body is the synthesis and release of vitellogenic proteins and lipids for yolk formation during oocytes maturation (Keeley, 1985; Kunkel and Nordin, 1985; Raikhel and Dhadialla, 1992). Sublethal doses of chlorfluazuron reduced the amounts of S. litura eggs constituents of protein, lipid and carbohydrates (Perveen, 2011). Soltani et al. (1996,1987) found that diflubenzuron affect both the weight and protein levels in the ovaries of T. molitor. Also affects the size of basal oocytes, the number of oocytes reproduction via the vitellogensis process. Zhu et al. (2012) found that treated S. litura with sublethal concentrations of hexaflumuron reduced larval survival and interfered with hemolymph physiological balances.
The aim of this work was to study the effect of sublethal dose of hexaflumuron on some biological aspects, ovary and egg maturation of the spiny bollworm, E. insulana.
MATERIALS AND METHODS
Insect rearing: The culture of spiny bollworm was obtained from Bollworms Research Department, Plant Protection Research Institute, Sharkia Brach. The emerged moths were sexed and each five pairs were confined in a glass jar (1/2 kg) covered with cloth as a suitable site for eggs deposition and kept under the constant conditions 26±1°C and 75±5% RH. Moths were fed on 10% honey solution absorbed on cotton piece that daily renewed. The clothes with the deposited eggs were transferred to a convenient glass jars. The newly hatched larvae were reared on modified artificial diet according to Rashad and Ammar (1985) and Amer et al. (2010).
Insecticide used: Hexaflumuron (Consult 10% EC) was supplied by Dow AgroScience.
Toxicological studies: Serial aqueous dilutions of hexaflumuron were prepared (100, 50, 25 and 12.5 ppm). Five grams of artificial diet were putted in each petri-dish and 1 mL of the tested concentration and water for the untreated was distributed on the surface of the diet and left until dryness. Each concentration and control was repeated five times. Twenty newly hatched larvae of the spiny bollworm transferred to the surface of the treated diet and untreated diet as control. Larvae were allowed to feed on the treated and untreated diet for one day after wards, live larvae were transferred individually to glass tubes (2×7 cm) containing untreated diet and incubated under the previous conditions. Larval mortality was recorded after four days. The LC values were calculated after four days according to Finney (1971).
Biological study: The newly hatched larvae were treated with LC20 as the previous method. Six replicates were used for treatment each replicate 50 larvae and the same were used for untreated. Larvae were allowed to feed on the treated diet for one day after wards, a live larvae were transferred individually to glass tubes (2×7 cm) containing untreated diet and incubated under the previous conditions until pupation. Newly emerged moths of the spiny bollworm were sexed and transferred to cages (five pairs/cage) and repeated three times. The moths were fed on 10% honey solution. Some biological aspects were recorded.
Ovary anatomy: Five female moths were dissected under stereomicroscope after emergence (0, 12, 24, 36 h), (2, 3, 4, 7, 10 days) and after post ovipostion. Female moths and dissected ovary were weighted. The ovary was separated and put on the stained trypan blue for 20 min at room temperature and excess stain was rinsed away with the saline 1.5%. Oocytes were defined as being mature and immature as staining by trypan blue response, mature eggs were unstained where immature stained dark blue (Telfer and Anderson, 1968; Webb et al., 1999). The number of mature and immature oocytes was counted using a binocular microscope (10×magnification) for the treated and untreated insects. The length and width of the basal oocytes and length of ovrioles were measured on the ovary of the newly emerged moths using micrometer slides. The number of spermatophores reception of the spiny bollworm female moths was counted after post ovipostion period.
Biochemical study: One gram from 12 days old larvae of the spiny bollworm were collected from treated and untreated for biochemical studies.
Total carbohydrates determination: Total carbohydrates were estimated in acid extract of insect by the phenol-sulphuric acid reaction (DuBois et al., 1956). Total carbohydrates were extracted and prepared for assay according to Crompton and Birt (1967).
Total lipids determination: Total lipids were estimated by the method of Knight et al. (1972) using phosphovanillin reagent prepared by dissolving of 0.6 gm pure vanillin in 10 mL ethanol and completed to 100 mL with distilled water. Then 400 mL conc. phosphoric acid was added. Two hundred and fifty microliter of sample were added to conc, sulphuric acid (5 mL) in a test tube and heated in a boiling water bath for 10 min. After cooling to room temperature, the digest was added to phosphovanillin reagent (6 mL). After 45 min, the developed color was measured at 525 nm against reagent blank, optical density was compared to that of a reference standard and results expressed as mg lipids/g body weight.
Determination of total soluble protein: Colorimetric determination of total soluble protein in supernatants of homogenate E. insulana larvae were carried out as described by Gornall et al. (1949) the principle of this method was based on the presence of an alkaline cupric sulfate, the protein produce a violet purple colour, the intensity of which is proportional to their concentration.
Data analysis: The data was analyzed using Costat program, COHORT software (CoStat, 2005).
RESULTS AND DISCUSSION
Effect of LC20 of hexaflumuron on some biological aspects: The LC20, LC50 and LC90 of hexaflumuron were 17.42, 72.75 and 1188.85 ppm, respectively. The slope value was 1.36±0.15 (Table 1). Accumulated larval mortality percentages were 28.56 and 3.66% for treated and untreated, respectively. Treatment of the newly hatched larvae with LC20 of hexaflumuron caused elongation of larval and pupal duration compared with untreated (Table 2). The average larval and pupal durations were 18.45 and 10.26 days, respectively for treated. While in untreated were 14.78 and 9.23 days, respectively. Hexaflumuron caused deformed larvae which failed to molt; the ecdysial cleavage was open but didn't completed and the larvae dead (Fig. 1e and f). The moths emergence percentages were 95.01 and 60.45% on untreated and treated, respectively. Hexaflumuron reduced the moths emergence significantly compared with untreated. Treatment insignificantly elongated female pre and post ovipostion compared with untreated, but shortened ovipostion period. The male longevity was 12.66 and 14.95 days for treated and untreated, respectively (Table 2). Treatment of newly hatched larvae with LC20 of hexaflumuron was reduced mean number of deposited eggs/female (85 eggs/female) compared with (180 eggs/female) for control (Table 3).
| Table 1: | Toxicity of hexaflumuron to newly hatched larvae of spiny bollworm |
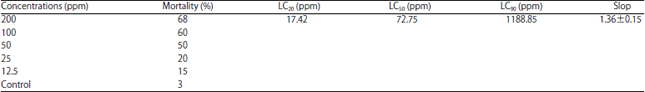 | |
| Table 2: | Effect of LC20 of hexaflumuron on some biological aspects of the spiny bollworm |
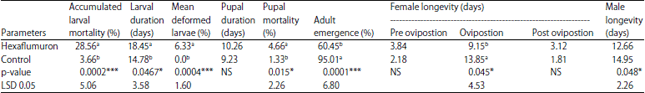 | |
| *Statistically significant as p<0.05, ***Highly significant as p<0.001 | |
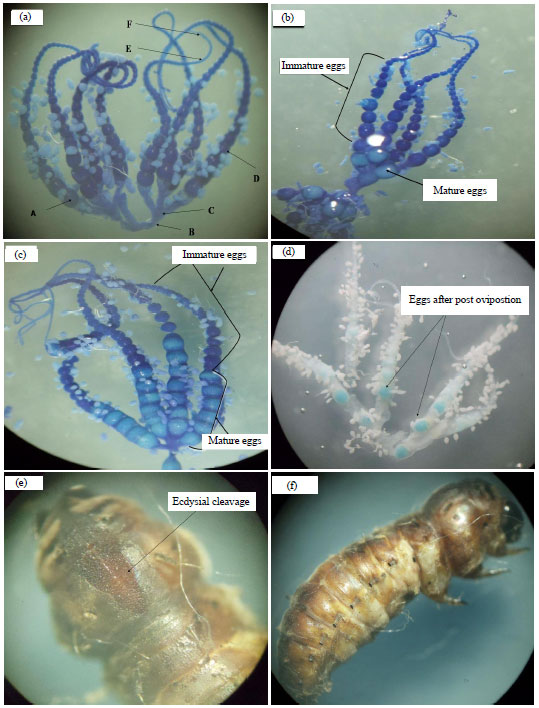 | |
| Fig. 1(a-f): | The stages of eggs maturation and deformed larvae, (a) A: Basel egg, B: Lateral oviduct, C: Calyx, D: Ovariol, E: Germarium, F: Terminal filament, (b) First appear of mature eggs after 36 h, (c) Peak of mature eggs after four days, (d) After post ovipostion period and (e, f) Deformed larvae failed to complete open ecdysial cleavage |
| Table 3: | Effect of LC20 of hexaflumuron on the spiny bollworm ovary, eggs and reproductive potential |
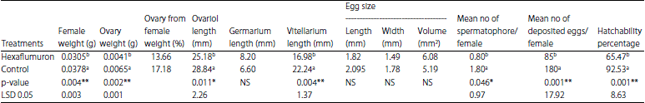 | |
| *Statistically significant as p<0.05, **Highly significant as p<0.01 | |
It may be resulted in effect of hexaflumuron on the ovariol length and reduced number of immature and mature eggs. Hatchability percentage was reduced significantly as affected by treated with hexaflumuron. The average hatchability percentages were 65.47 and 92.53% for treated and untreated, respectively, the decreased of hatchability percentage may be due to decrease number of spermatophor/female in treatment.
Weight of female and ovary: Treated the newly hatched larvae with hexaflumuron was caused significant reduced in the newly emerged female moths weight compared with untreated. The averages weight of female moths were 0.0378 and 0.0305 g/female moth on untreated and treated, respectively. Also, weight of ovary was significantly reduced on treated compared with untreated. The average weights of ovary were 0.0065 and 0.0041 g/ovary, respectively. The ovary consists of 13.66 and 17.18 % from the weight of moth, respectively (Table 3).
Ovariol length: In newly emerged moth resulted from treated larvae with LC20, the length of ovariole was reduced insignificantly to that of untreated (Table 3). The lengths were 28.84 and 25.18 for untreated and treated, respectively. The treated was caused elongation on the germarium compared with untreated.
Size of basal oocyte: Length and width of the basal oocytes were measured on the ovary of the newly emerged moths. Treated was reduced insignificantly size of the basal eggs compared with untreated. The size of basal oocyte was 6.08 and 5.19 mm2 on untreated and treated, respectively (Table 3). Treated newly hatched larvae with LC20 was reduced the size of basal eggs with 14.63% on the ovary of the newly emerged moths.
Number of spermatophores: The average number of spermatophores reception of the spiny bollworm female moths were 0.80 and 1.80 spermatophores/ female for treated and untreated, respectively (Table 3). Treatment significantly reduced number of female mated compared with those of control.
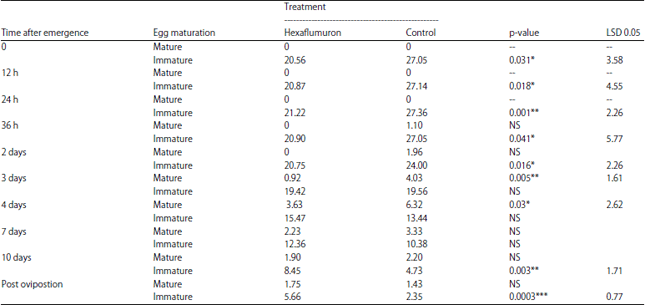
Egg maturation: The first appearance of mature eggs in the female moths ovarioles was took place after a period of 36 h from emergence in untreated and three days in treated with hexaflumuron (Table 4). The averages mature eggs were 1.10 and 0.92 eggs/ ovariol after 36 h and three days, respectively. The average egg maturation was increased gradually until the 4th days after moths emergence in treated and untreated. After that the number of mature eggs was decreased until ten days from emergence. Earias insulana larvae fed on LC20 of hexaflumuron had a decrease in the numbers of egg formations in the ovarioles as compared with untreated. Means of 216.40 and 164.48 immature oocytes were recorded in moths after emergence in untreated and treated, respectively (Fig. 1a-d).
| Table 4: | Effect of LC20 of hexaflumuron on the spiny bollworm egg maturation |
 | |
| *Statistically significants as p<0.05, **Highly significant as p<0.01, ***Highly significant as p<0.001 | |
| Table 5: | Effect LC20 of hexaflumuron on total carbohydrate, lipid and soluble protein on spiny bollworm larvae |
 | |
| *Statistically significant as p<0.05, **Highly significant as p<0.01 | |
The average numbers of immature eggs were found in the ovary after ovipostion period was significant increased on treated moths compared with those untreated (Table 4).
The average numbers of eggs found after post ovipostion were 30.24 and 59.28 eggs/moths on untreated and treated, respectively. The treated newly hatched larvae of the spiny bollworm with LC20 of hexaflumuron reducing the number of oocyte formation and the mature eggs and increased eggs on the ovary after ovipostion period compared with untreated.
Effect of hexaflumuron on some biochemical parameters on E. insulana larvae: The mean total carbohydrate, lipid and soluble protein in larvae treated with hexaflumuron showed a significant decreased compared with untreated (Table 5). The means were 7.99, 136.68 and 18.22 mg g–1 for treated compared with 4.98, 104.37 and 12.49 mg g–1 for control, respectively.
DISCUSSION
Treated the newly hatched larvae of the spiny bollworm with LC20 of hexaflumuron caused elongation larval and pupal duration, decreased moths emergence percentage and weight of larval. Mahmoudvand et al. (2011) found that sublethal concentrations of hexaflumuron reduced fecundity of Plutella xylostella females significantly but hatchability of treatments and control were similar. The LC50 values were 0.31 and 0.46 mg ai/L in dietary method and leaf disc method, respectively against 1st instar larvae of Helicoverpa armegera (Dastjerdi et al., 2008). Zhu et al. (2012) found that treated S. litura with sublethal concentrations of hexaflumuron reduced larval survival and interfered with hemolymph physiological balances. Treatment of hexaflumuron reduced number of eggs on the ovary after emergence, decreased number of immature and mature eggs, size of basal oocyte and length of ovarioles and number of deposited eggs and hatchability percentage compared with untreated. Treated larvae reduced the larval content of total carbohydrate, lipid and soluble protein. Reduced the number of eggs on the spiny bollworm ovary after female emergence and the eggs maturation may be due to reduce the larval content of lipid and protein as affected by treated the larvae by LC20 of hexaflumuron (El-Nemaky and Azab, 2004). Results showed that hexaflumuron was active at 0.5 and 0.25 ppm against one-day-old larvae of E. insulana. Horowitz et al. (1992) found that the LC50 of hexaflumuron ingested by newly hatched larvae of the spiny bollworm was 0.25 ppm. Perveen and Miyata (2000) found that treated the fifth instar larvae of S. litura with sublethal doses of chlorfluazuron (1.00 ng/larva or 3.75 ng/larva) topically reduced ovarian weight and number of mature eggs in pupae and adults compared with those of control. Also, disrupt growth and development of oocytes by significant affecting the size of basal oocytes. Protein content of ovarian was significant reduced. Soltani and Soltani-Mazouni (1997) studied diflubenzuron topically application to C. pomonella at pupal ecdysis disturbed growth and development of oocytes. Treatment reduced size of basal oocytes and the number of oocytes per ovary in newly emerged adults. Soltani et al. (1996, 1987) found that diflubenzuron affect both the weight and protein levels in the ovaries of T. molitor. Also affects the size of basal oocytes, the number of oocytes reproduction via the vitellogensis process. Khebbeb et al. (1997) found that diflubenzuron quantitatively and qualitatively change the lipid metabolism during sexual maturation in T. molitor.
CONCLUSION
Used hexaflumuron to control cotton pests was caused reduction on the deposited number and hatchability percentage of eggs in adding to reduce moths emergence of E. insulana.
REFERENCES
- Crompton, M. and L.M. Birt, 1967. Changes in the amounts of carbohydrates, phosphagen and related compounds during the metamorphosis of the blowfly, Lucilia cuprina. J. Insect Physiol., 13: 1575-1592.
CrossRefDirect Link - Dastjerdi, H.R., M.J. Hejazi, G.N. Ganbalani and M. Sabe, 2008. Toxicity of some biorational and conventional insecticides to cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae) and its ectoparasitoid, Habrobracon hebetor (Hymenoptera: Braconidae). J. Entomol. Soc. Iran, 28: 27-37.
Direct Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Gornall, A.G., C.J. Bardawill and M.M. David, 1949. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem., 177: 751-766.
CrossRefPubMedDirect Link - Horowitz, A.R., M. Klein, S. Yablonski and I. Ishaaya, 1992. Evaluation of benzoylphenyl ureas for controlling the spiny bollworm, Earias insulana (Boisd.), in cotton. Crop Protect., 11: 465-469.
CrossRefDirect Link - Khebbeb, M.E.H., J. Delachambre and N. Soltani, 1997. Lipid metabolism during the sexual maturation of the mealworm (Tenebrio molitor): Effect of ingested diflubenzuron. Pestic. Biochem. Physiol., 58: 209-217.
CrossRefDirect Link - Knight, J.A., S. Anderson and J.M. Rawle, 1972. Chemical basis of the sulfo-phospho-vanillin reaction for estimating total serum lipids. Clin. Chem., 18: 199-202.
CrossRefPubMedDirect Link - Mahmoudvand, M., H. Abbasipour, A.S. Garjan and A.R. Bandani, 2011. Sublethal effects of hexaflumuron on development and reproduction of the diamondback moth, Plutella xylostella (Lepidoptera: Yponomeutidae). Insect Sci., 18: 689-696.
CrossRefDirect Link - Perveen, F., 2011. Effects of sublethal doses of chlorfluazuron on ovarioles in the common cutworm, Spodoptera litura (F.) (Lepidoptera: Noctuidae). J. Life Sci., 5: 609-613.
Direct Link - Perveen, F. and T. Miyata, 2000. Effects of sublethal dose of chlorfluazuron on ovarian development and oogenesis in the common cutworm Spodoptera litura (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am., 93: 1131-1137.
Direct Link - Raikhel, A.S. and T.S. Dhadialla, 1992. Accumulation of yolk proteins in insect oocytes. Annu. Rev. Entomol., 37: 217-251.
CrossRefDirect Link - Soltani, N. and N. Soltani-Mazouni, 1997. Oogenesis in mealworms: Cell density of Germarium, thickness of chorion and ecdysteroid production. Effects of regulators. Med. Fac. Landbouww. Univ. Gent., 62: 565-571.
Direct Link - Soltani, N., A. Quennedey, J.P. Delbecque and J. Delachambre, 1987. Diflubenzuron-induced alterations during in vitro development of Tenebrio molitor pupal integument. Arch. Insect Biochem. Physiol., 5: 201-209.
CrossRefDirect Link - Soltani, N., N. Soltani-Mazouni and J. Delachambre, 1996. Evaluation of Triflumuron, a benzoylphenylurea derivative, on Tenebrio molitor pupae (Col., Tenebrionidae): Effects on cuticle. J. Applied Entomol., 120: 627-629.
CrossRefDirect Link - Telfer, W.H. and L.M. Anderson, 1968. Functional transformations accompanying the initiation of a terminal growth phase in the cecropia moth oocyte. Dev. Biol., 17: 512-535.
CrossRefDirect Link - Webb, T.J., S. Shu, S.B. Ramaswamy and S. Dorn, 1999. Influence of juvenile hormone and mating on oogenesis and oviposition in the codling moth, Cydia pomonella. Arch. Insect Biochem. Physiol., 41: 186-200.
CrossRefPubMedDirect Link - Amer, A.E.A., A.A.A. El-Sayed and S.A.A. Raslan, 2010. Improved techniques for laboratory rearing of the spiny bollworm, Earias insulana (Boisduval) (Lepidoptera: Noctuidae). J. Plant Protect. Pathol. Mansoura Univ., 1: 299-306.
Direct Link - Zhu, Q., Y. He, J. Yao, Y. Liu, L. Tao and Q. Huang, 2012. Effects of sublethal concentrations of the chitin synthesis inhibitor, hexaflumuron, on the development and hemolymph physiology of the cutworm, Spodoptera litura. J. Insect Sci., 12: 1-13.
CrossRefPubMedDirect Link