
Amany M.H. Abu-shall
Department of Applied Entomology, Faculty of Agriculture, Alexandria University, Egypt
M.E. Tawfeek
Department of Applied Entomology, Faculty of Agriculture, Alexandria University, Egypt
Journal of Entomology
Year: 2015 | Volume: 12 | Issue: 2 | Page No.: 67-76







ABSTRACT
The lycaenid butterfly Chilades pandava Horsfield (Lepidoptera: Lycaenidae) is an important pest of many species of cycas trees. It is recorded for the first time in Egypt in 2013. The adults were collected from the infested cycas trees in arboretum of Faculty of Agriculture, Alexandria University. Each part of male and female were dissected and prepared as whole mounts for light and SEM microscopic examination. Morphology of the cycas blue butterfly C. pandava (Lepidoptera: Lycaenidae), adult and the ultrastructure of its antennal sensilla were described and illustrated by SEM photographs.
PDF Abstract XML References Citation
Received: January 21, 2015;
Accepted: April 14, 2015;
Published: August 25, 2015
How to cite this article
Amany M.H. Abu-shall and M.E. Tawfeek, 2015. Description of the Egyptian Form of Chilades pandava Horsfield (Lepidoptera: Lycaenidae: Polyommatinae) and Ultrastructure of Antennal Sensilla. Journal of Entomology, 12: 67-76.
DOI: 10.3923/je.2015.67.76
URL: https://scialert.net/abstract/?doi=je.2015.67.76
DOI: 10.3923/je.2015.67.76
URL: https://scialert.net/abstract/?doi=je.2015.67.76
INTRODUCTION
The lycaenid butterfly Chilades pandava Horsfield (Lepidoptera: Lycaenidae) is an important pest of many species of cycas trees (Chang, 1989; Marler et al., 2012). It is recorded for the first time in Egypt (2013) and the African continent by Fric et al. (2014). Kunte and Tiple (2009) mentioned that many butterflies species show environmentally induced seasonal forms, known as seasonal polyphenism. They added that there is geographic variation in the occurrence of seasonal forms in C. pandava. For example the seasonal forms have not been reported in Seri Lanka subspecies or in that in the humid tropical areas in south-east Asia. Therefor, the aim of this study is to describe the Egyptian form of the cycas butterfly, male and female, since it is newly recorded pest in the Egyptian fauna, to help the worker in horticulture and economic entomology fields to distinguish this destructive pest.
The antenna of insects have various types of sensilla that play important roles in insect behavior, including habitat searching, host recognition, feeding, mate attraction and ovipositon. It is generally agreed that the sensilla on insect antennae are not randomly distributed (Zacharuk, 1985; Skiri et al., 2005). The various types of sensilla function as tactile, smell receptors, contact chemoreceptors, hygroreceptors and temperature receptors. This study, also deals with studying the fine structure of the antennal sensory receptors as an attempt to provide basic information for future of electrophysiological work concerned to the behavior of this pest.
MATERIALS AND METHODS
Collections and preparations
Insect collections: Chilades pandava (Lepidoptera: Lycaenidae: Polyommatinae) adult were collected from the infested cycas trees in arboretum of Faculty of Agriculture, Alexandria University.
Light microscope preparations: Each part of male and female were dissected and prepared as whole mounts for light microscopic examination by soaking in 10% NaOH solution for a week then rinsed in distilled water several times. The specimens passed through series of ethyl alcohol from 60-95% then to absolute alcohol 1 h for each concentration. After that they were cleared in clove oil for 1 h and then mounted on slides using Canada balsam medium. The slides were dried at 50°C. For wing description, the terminology of Kunte and Tiple (2009) was used (Fig. 1). The nomenclature and terminology of genitalia is according to Sibatanp et al. (1954).
The veins are numbered. The wing pattern elements demarcated by veins are organized into several series of spots, some of which form bands: (1) Cell-end band; (2) Discal band; (3) Submarginal band; (4) Costal spots in the wing space 7; (5) Sub-basal spots in space 7, followed by 3 sub basal spot in the cell and space 1c and 1a and (6) Tornal orange-crowned black spots (Kunte and Tiple, 2009).
Scanning Electron Microscopy (SEM) preparations: For Scanning Electron Microscopy (SEM) examination the antennae were dried in a series of ethanol to 96% then mounted in a stub by means of double face sticky transparent tape. The specimens were coated with gold in (JOELFC-1100E) high resolution sputter coater for 25 min to a thickness of 12 nm. Preparations were examined in JOEL (JSM 5300) Scanning Electron Microscopy at the Faculty of Science, Alexandria University.
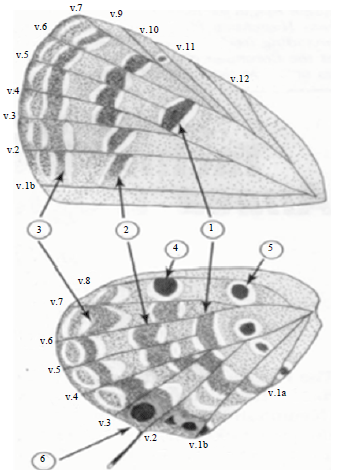 | |
| Fig. 1: | Wing pattern elements of C. panadava |
RESULTS AND DISCUSSION
Description of adult: Male is about 10-13 mm (11.5 mm in average) in length. The wing span is about 21-23 mm (22 m in average). Female slightly longer averaged 13 mm in length and wing span from 27-29 mm (28 m average).
This insect is violet-blue or lavender-blue in colour in both sexes but the female slightly paler with shiny blue scales on the basal half of wings. Body is blue dorsally and light brown ventrally, antenna black with white rings.
Head: The large compound eye with short erect hairs between facets. Simple eyes are absent. The flagellum of the clavate antenna consists of 17 filiform segments followed by 17 clubbed ones in both sexes. Antennae covered with dense overlapping scales (Fig. 2) and provided with different types of sensilla. Mouth parts consist of coiled proboscis provided with sense organs and a pair of three-segmented labial palps.
Thorax: The prothorax is the smallest of the three thoracic segments, with a pair of spiracles.
Female legs are similar in shape and size with five segmented tarsus, bearing a pair of forked claw (Fig. 3a, b) while fore leg in male is reduced with only one segmented tarsus ends with a single claw (Fig. 3c).
Female wings: The upper side (Fig. 4a) of the fore wing is shiny metallic blue in colour, costa and marginal broad, dusky black. Thin antiapical black line followed by grayish cilia at terminal edge. Hind wing broader than front wing, metallic bluish in colour, costa slightly broad, dusky black. The submarginal area has eight blackish spots each surrounding with a grey ring. Six of them are large and 2 small. First and second ones near apical angle are obsolescent, the next four are large. Spot number 6 capped with orange crown. Hind wing with blackish filamentous tail.
 | |
| Fig. 2: | SEM photograph showing dense overlapping scales covered antennae of Chilades pandava |
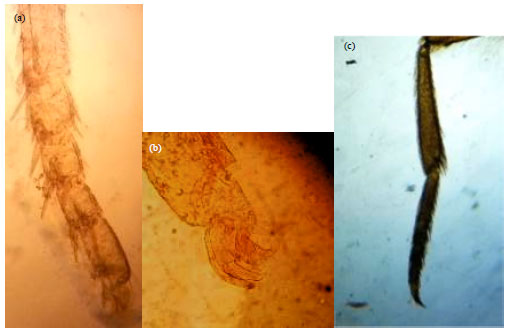 | |
| Fig. 3(a-c): | End of fore leg, (a) Female, (b) Tarsal claw of female fore leg and (c) Male |
The underside of fore wing is fuscous light brown in colour with brown markings. There is a median transverse brown rectangular band (cell end band) with white sides. Discal band consists of a series of brown spots with thin white sides followed by a sinuate brownish, whitish band. The submarginal area distinguished into an inner brown band and an outer 6 brown oval spots surrounding by lighter whitish colour. Termine is brown thin line.
Hind wing underside has a dark brown bar at the terminal end of the discal cell. The postmedian area with a dark brown discal band white sided, composed of spots that are bounded by wing veins. The subterminal area distinguished into an inner brown band and an outer one consists of a series of spots, the four tornal spots small, brown with white boarder, followed by large black spot crowned with orange cap, just above the tail. The next four large are brown with white ring. Termine is brown thin line just before the light brown cilia.
Male wing: Upper side (Fig. 4b) of male fore wing blue with thin costa and terminal margin fuscous brown. The terminal edge with anticiliary black line and Cilia dark brown. Hind wing costa narrowly fuscous brown, subterminal area with a series of black spots edged with white circle. The spot in space 2 is the largest and crowned with orange cap anterior to the black wing tail. The anticiliary and cilia as fore wing. Under side of hind wing as in the female.
It is worth to mention that seasonal variations were observed in the appearance of wings colour during the present study which are not in our consideration in this investigation.
Abdomen: It comprises 10 segments, the first seven having spiracles. The last two or three segments are greatly modified to form the external genitalia (Braby, 2000). There is a dense tuft of long hairs (pencil hairs) at the end of male abdomen associated to scent gland and used for the courtship (Fig. 5).
 | |
| Fig. 4(a-c): | Upper side of Chilades pandava (a) Female, (b) Male and (c) Under side of both male and female |
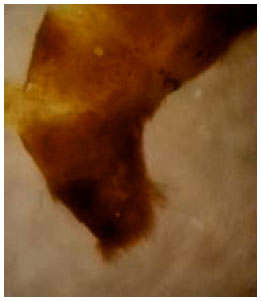 | |
| Fig. 5: | Hair pencil of male |
Male genitalia: The external male genitalia is formed by the appendages of the 9th and 10th abdominal segments. The ninth segment forms a sclerotized dorsal tegumen and a ventral vinculum. Sacus is a process found midventrally forwards from the vinculum while uncus is a pair curved down middorsal attached posteriorly to the tegumen. A pair of lateral valve or claspers protruded below vinculum.
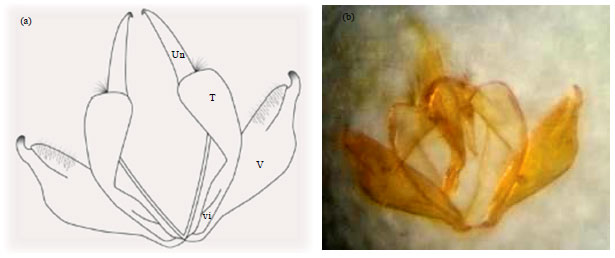 | |
| Fig. 6(a-b): | Chilades pandava male genitalia (a) Diagram and (b) Photograph, T: Tegument, Un: Uncus, V: Valva, Vi: Vinculum |
 | |
| Fig. 7: | Bursa copulatrix of Chilades pandava female genitalia |
Gnathos is a pair of long separate hooks known as brachia attached also posteriorly to the tegumen. The aegaeus is a centrally sclerotized hollow tube through the transverse membrane between the bases of claspers (Fig. 6).
Female genital: As in most lepidopterous adult females have two genital openings, one at the posterior end of the abdomen, used for egg laying and the other, on the midventral line between the 7th and 8th sternites, is used for mating. Segments 8, 9 and 10 form a retractile ovipositor. A pair of hairy anal pads, are found at the end of a pair of internal rod-like apophysis. The copulatory opening is bordered by sclerotized plates and leading to a long bursa duct, the latter leading to the membranous sac (corpus bursa) in which the spermatophores were put. The internal surface of the corpus bursa supported with two thickened and rough plates, known as signa (Fig. 7).
 | |
| Fig. 8: | SEM photograph showing the Bohm’s bristles (Bb.) of C. pandava antennae |
Ultrastructure of antennal sensilla: This part of study dealing with the different types of the antennal sensilla or receptors of adult Chilades pandava Horsfield which was studied by Scanning Electron Microscopy (SEM). There was five types of sensillae in both sexes as follows.
Böhm’s bristles (Bb.): They are a group of short spin-like sensilla, about 5 μm in length, with smooth wall, found on both sides of bases of scape and pedicel. According to Schneider (1964), Bohm’s bristles are present on homologous places in all lepidopterous insects, this strategic places at articulations indicated that they may be considered as mechanoreceptors. They function as proprioceptors perceiving the antennal position. On the other hand, Callahan et al. (1968) mentioned that these tapered configuration, extremely narrow walls, heavy socket-mounted base, dome sensors might operate as light waveguide aerials in the visible region and function as a gating system for the response of moth antenna to parameter such as scent, temperature and moisture in the surrounding environment (Fig. 8).
Sensilla chaetica (Sc.): It is about 10.7 μm in length, straight spine-like sensilla arising from a rounded socket with radial ridges. It is the only type of sensilla which found on one side the cylindrical segments of antennae. They are arranged on one row (14 setae) on the clavat flagellomeres among the trichoid sensilla except the distal flagellomere which has two rows of them (Fig. 9 and 10a).
Sensilla trichoidia (St.): They are numerous sensilla on all flagellomeres. They are slender, curved or nearly straight, thin walled hair distally, tapering and inserted in socket. It is relatively long about 15.65 μm in length and found in antennae of many insects (Schneider, 1964) (Fig. 9 and 11).
Sensillae ceoloconica (Sco.): They are found in a circular tubercle about 5.71 μm in diameter, there are 3 tubercles in male and two in female at the distal part of last segment of antennae.
 | |
| Fig. 9: | SEM photograph of C. pandava antennae showing Sc: Sensilla chaetica, Sa: Stellate sensilla, St: Sensilla trichoidea |
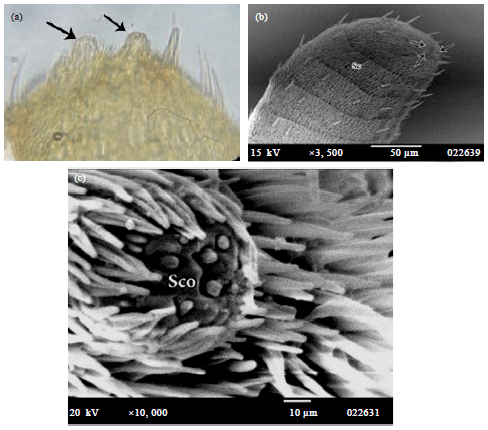 | |
| Fig. 10(a-c): | Chilades pandava antennae showing (a) Microscopic photo of distal flagellomere, (b) SEM photograph of the clavate flagellomeres and (c) Magnification of circular tubercle which bearing sensilla coeloconic (Sco.). Arrows indicated place of tubercles on the apex of last antennal segment, Sc: Sensilla chaetica |
 | |
| Fig. 11: | SEM photograph showing Sa: Sensilla stellate and St: Sensilla trichodea of C. pandava antennae |
Each tubercle is surrounded by many sensilla and bearing nine sensilla ceoloconica about 0.71 μm in length. The latter, are thin-walled doom shaped sensilla on the floor of depression (pit about 1 μm in diameter) (Fig. 11).
Stellate sensilla (Sa.): Sensilla stellate are cone shaped, with longitudinal deep ridges. They are numerous, distributed along the clavate flagellomeres .
Numbers of antennal sensilla were different between female and male adults but the category, shape and distribution were the same between them (Jian et al., 2011) also, found 5 types of C. pandava antennal sensilla, sensilla stellate, sensilla trichodea, sensilla chaetica, sensilla basiconica and Böhm bristles were on antennae. The present study agrees with the finding of hem but disagree with the presence of sensilla basiconica (Fig. 11).
CONCLUSION
Morphology of the cycas blue butterfly Chilades pandava (Lepidoptera: Lycaenidae), adult and the ultrastructure of its antennal sensilla were described and illustrated by SEM photographs.
ACKNOWLEDGMENT
The authors wish to express their sincere thanks and gratitude to Dr. Hanan M. Ramadan, Professor of Economic Entomology, Faculty of Agriculture, Alexandria University for her valuable technical help and advice throughout the achievement of this study and to Dr. Hedaya H. Karam Professor in the same Department for reviewing the manuscript.
REFERENCES
- Fric, Z., R. Dickinson, G. Fetouh, T.B. Larsen, W. Schon and M. Wiemers, 2014. First record of the cycad blue, Chilades pandava, in Egypt—A new invasive butterfly species in the Mediterranean region and on the African continent (Lepidoptera: Lycaenidae). Afr. Entomol., 22: 315-319.
CrossRefDirect Link - Kunte, K. and A. Tiple, 2009. The polyommatine wing pattern elements and seasonal polyphenism of the Indian Chilades pandava butterfly (Lepidoptera: Lycaenidae). News Lepidopterists Soc., 51: 86-109.
Direct Link - Marler, T.E., A.J. Lindstrom and L.I. Terry, 2012. Chilades pandava damage among 85 Cycas species in a common garden setting. HortScience, 47: 1832-1836.
CrossRefDirect Link - Jian, M.L., L.L. Zhang and R.Q. Mao, 2011. Studies on the antennal sensilla of Chilades pandava by scanning electron microscopy. J. South China Agric. Univ., 32: 52-56.
Direct Link - Skiri, H.T., M. Stranden, J.C. Sandoz, R. Menzel and H. Mustaparta, 2005. Associative learning of plant odorants activating the same or different receptor neurones in the moth Heliothis virescens. J. Exp. Biol., 208: 787-796.
PubMed