
Anar A. Bakr
Depertment of Applied Entomology, Faculty of Agriculture, (Elshatby) Alexandria University, Alexandria, Egypt
Journal of Entomology
Year: 2013 | Volume: 10 | Issue: 1 | Page No.: 43-48







ABSTRACT
Adults and newly laid eggs of the astigmatid mite, Tyrophagus putrescentiae were exposed to UV-C (260 nm) and UV-B (315 nm). In adults, the most effective treatments were observed from distance 10 cm and exposure times ranged between 3-15 min with mortality rates between 85-100% for UV-C and 62-83% for UV-B. Based on the LD50 values the UV-C radiation was more effective than the UV-B at different exposure times within each distance tested (5, 10 and 20 cm). A positive correlation was detected between the UV dose and mortality within each distance used for UV-C and UV-B irradiances. Regarding eggs none of the UV-B or UV-C exposed eggs in all treatments hatched.
PDF Abstract XML References Citation
Received: September 09, 2012;
Accepted: November 20, 2012;
Published: January 28, 2013
How to cite this article
Anar A. Bakr, 2013. Effectiveness of Ultraviolet Radiation as a Physical Method in Controlling the Stored Product Mite, Tyrophagus putrescentiae (Acari: Acaridae). Journal of Entomology, 10: 43-48.
DOI: 10.3923/je.2013.43.48
URL: https://scialert.net/abstract/?doi=je.2013.43.48
DOI: 10.3923/je.2013.43.48
URL: https://scialert.net/abstract/?doi=je.2013.43.48
INTRODUCTION
Over the last few decades, the diets of humans have changed and now include more food grains and stored foods with high fat and protein content. These changes are conducive to the growth and cosmopolitan occurrence of stored-food mites. The astigmatid mite, Tyrophagus putrescentiae (Schrank), is one of the most important storage mites and causes serious economic losses (Zdarkova, 1991) as well as a reduction of nutrient contents and seed viability (Krantz, 1955). In addition, the mite is known to be the etiological agent of allergic diseases among farmers and workers handling heavily infested stored products (Hughes, 1976). Moreover, systemic anaphylaxis was reported from eating T. putrescentiae infested food (Matsumoto et al., 1996). Mite also acts as a carrier of bacteria and toxigenic fungi (Aspergillus spp. and Penicillium spp.) in stored grain kept under warm and moist conditions (Van Bronswijk and Sinha, 1973; Franzolin et al., 1999).
Accordingly, various attempts to control these mite populations have been implemented using chemical methods. Changes in public attitudes towards the use of chemicals to control pest have resulted from increased concern for food safety and preservation of environmental quality. Therefore, there is a need for non-chemical, non-residue method of efficiently and effectively destroying mites in food supplies.
Artificial Ultraviolet (UV) irradiation could prove to be a promising rapid physical method that satisfies that need. The range of UV wavelength is often subdivided into UV-A (400-315 nm); UV-B (315-280 nm) and UV-C (280-200 nm), UV-C also called short wave or germicidal wavelength.
UV-B can directly damage DNA and is a strong oxidative stressor (Tan et al., 2007), so it can be expected to be lethal to small organisms such as mites. Also, UV-C ray can irradiate many organisms like bacteria, viruses and mites as it has enough energy to damage chemical bonds in DNA and RNA molecules (Goodsell, 2001). The ultraviolet light is absorbed by surface colouring chemicals in the mite, such as integument, which act as a heat sink to the ultraviolet photons. In this manner, the undesired mites are selectively heated without harming adjacent food objects.
The present study investigates the susceptibility of T. putrescentiae adults and their newly laid eggs to UV-B and UV-C radiation at different exposure times from different distances.
MATERIALS AND METHODS
Laboratory culture of T. putrescentiae: A stock culture of T. putrescentiae was established from infested samples from Alexandria grain stores. The respective cultures were maintained without exposure to any known acaricide. The mites were reared on a standard mite diet of dried yeast and whole meal flour (3:1) in glass containers (10 cm diameter and 12 cm height), each covered with a round plastic plate having a 3 cm diameter hole in the center which was sealed with a filter paper disk for ventilation (Sanchez-Ramos and Castanera, 2001). The rearing containers were kept at 25±1°C and 75±5 relative humidity (Jeong et al., 2009). Stages which were subjected to the experimentation placed into a glass microcell (1.5 cm diameter and 2 cm depth) covered with a lid.
UV treatment
Effects on adults: The effects of UV-B and UV-C radiations on the mortality in T. putrescentiae female adults (7-10 days old) were determined by using the monochromatic photons. Selected monochromatic photons of UV light was generated from two sources, a 15 watt Black light FL 8 BL lamp with wavelength (315 nm) for UV-B and a G15 T8/15 watt lamp with wave length (260 nm) for UV-C. The lamps were suspended above and perpendicular to the microcells into metal boxes. In each glass microcell placed in the boxes with radiation wavelengths of 260 and 315 nm, T. putrescentiae adults that were placed beneath a mirror that reflected the monochromatic light beam directly onto them. Table 1 lists the distance, exposure time and dosage at 15 W irradiance from UV-B and UV-C lamps. Treated and control mites were held at the same conditions used for colony maintenance. The number of T. putrescentiae female adult that were alive or dead were observed 24 h after irradiation periods with a stereomicroscope.
Effect on eggs: T. putrescentiae eggs laid within 24 h were placed as described above. The maximum exposure time previously used with each distance was tested, since eggs usually need higher doses than mobile stages. Egg development was checked daily under a stereomicroscope and hatchability was monitored for 10 days. Specimens were confined at optimum conditions and compared to untreated controls.
Statistical analysis: Statistical significant differences between mortality incidence under control and those exposed to different exposure times at a specific distance were determined by Duncan’s multiple range test. Correlation analysis was conducted to determine UV dose response in mortality.
| Table 1: | Distance, exposure times and integrated doses tested on T. putrescentiae female adults |
 | |
RESULTS AND DISCUSSION
Mortality: In the UV-B treatments from 5 cm distance (Table 2), differences between mortality at the longest durations (30 sec and 2 min) and those at the lowest ones (20 and 10 sec) were significant. The greatest mortality rate (58%) was achieved at the longest exposure time (2 min), while the lowest mortality (35%) was detected with the least exposure time (10 sec). With the UV-C irradiance the best result was recorded at 2 min exposure time (69% mortality). Nearly mortality percentages were observed with UV-C irradiance at the four exposure times with no significant differences between them and those under UV-B irradiance (p>0.05).
Mortality percentages from 10 cm distance (Table 3) in all UV treatments of UV-C were significantly higher than those under UV-B irradiance at 3, 5 and 15 min (p>0.05) and 10 min (p<0.01). Interestingly, under UV-C irradiance full mortality (100%) was observed after 15 min. A mortality range of 85-97% was detected with exposure time ranging between 3 to 10 min. On the other hand, under the same range of exposure time with UV-B irradiance, mortality ranged between 62-67%. After 15 min of exposure time, mortality ranked the highest grade with a percentage 83%.
Table 4 shows that adults of T. putrescentiae from 20 cm distance under UV-C irradiance are significantly susceptible to all the tested exposure times than those under UV-B irradiance (p<0.01). The highest mortality rate was recorded with 30 min exposure time either for UV-B or UV-C irradiance, recording 52 and 85% mortality, respectively. while, the lowest mortality percentage was observed with the least exposure time (10 min) for either UV-B or UV-C irradiances, recording 35 and 69% mortality, respectively.
| Table 2: | Mortality in T. putrescentiae adults exposed to UV-B and UV-C treatments from 5 cm distance at four exposure times |
 | |
| Values (Means±SE) followed by the same letter in each column are not significantly different at p>0.05 | |
| Table 3: | Mortality in T. putrescentiae adults exposed to UV-B and UV-C treatments from 10 cm distance at four exposure times |
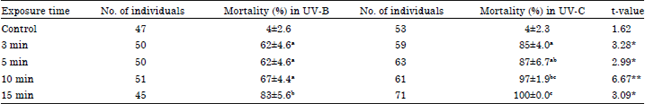 | |
| Values (Means±SE) followed by the same letter in each column are not significantly different at p>0.05, *p<0.05 and **p<0.01 | |
| Table 4: | Mortality in T. putrescentiae adults exposed to UV-B and UV-C treatments from 20 cm distance at four exposure times |
 | |
| Values (Means±SE) followed by the same letter in each column are not significantly different at p>0.05, *p<0.05 and **p<0.01 | |
With the adults of T. putrescentiae, it was observed that UV-B (315 nm) and UV-C (260 nm) in certain doses promoted mortality (Table 2-4). Suzuki et al. (2009) also showed that UV-C (250 nm) and UV-B (300 nm) increased mortality and escape and inhibited oviposition in non-diapausing females of T. urticae. Ultraviolet induces the hemolysis of hydrogen peroxide (H2O2) into hydroxyl radical, which seems to be the most damaging of the free radicals (Tan et al., 2007). UV radiation is observed by some coenzymes and pigments, raises these compounds to an excited state and the excitation energy is finally transferred to H2O molecules, yielding Reactive Oxygen Species (ROS). It has been suggested that UV-induced ROS damages important intra- and extracellular components, such as lipids, lipid membranes, nucleic acids and proteins especially when the duration of exposure to UV light is sufficient (Jurkiewicz and Buettner, 1994; Shindo et al., 1994; Goodsell, 2001). Such UV/ROS-induced damage would be lethal to mites because of their small size (their body surface area per weight is large).
Dose-response curves: A significant positive correlation was detected between the UV dose and mortality incidences in T. putrescentiae adults within each distance used. From 5 cm distance, the correlation values were 0.8317 and 0.5655 in the UV-B and UV-C, respectively (Fig. 1a). These values were 0.949 in the UV-B and 0.8791 in the UV-C from 10 cm distance (Fig. 1b), while from 20 cm distance they were 0.9287 and 0.9426 in the UV-B and UV-C, respectively (Fig. 1c).
In the present study, the above-mentioned effects of UV-C were stronger than those of UV-B when compared within each distance tested. Based on the 24 h LD50 values against T. putrescentiae adults, UV-C was approximately 1.8 (LD50 = 0.39 kJ cm-2), 1.7 (0.95 kJ cm-2) and 6.9 (4.15 kJ cm-2) times more effective than UV-B (LD50 = 0.72, 1.67 and 28.83 kJ cm-2) from 5, 10 and 20 cm distances, respectively (Fig. 1).
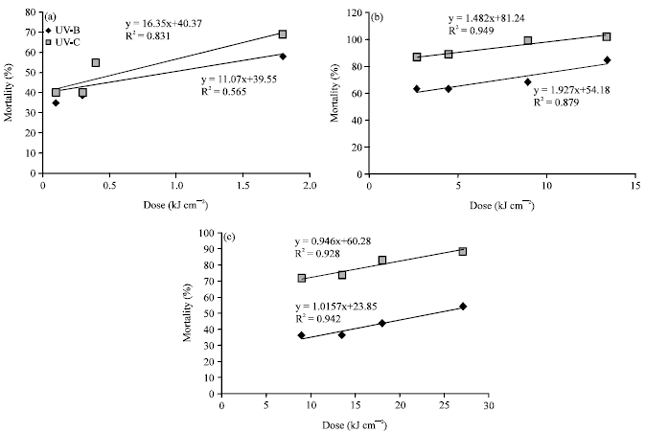 | |
| Fig. 1(a-c): | Dose-response relationship between the UV dose and mortality percentage of T. putrescentiae adults from 3 distances (a) 5 cm, (b) 10 cm and (c) 20 cm |
| Table 5: | Cumulative hatchability (%) for 10 days of T. putrescentiae eggs exposed to UV-B and UV-C radiations from three distances at three exposure times |
 | |
These results are relevant to those obtained by Suzuki et al. (2009). who reported that the median effective doses for 50% mortality plus escape (ED50) were 21 (UV-C) and 104 kJ cm-2 (UV-B) in non-diapausing females of Tetranychus urticae.
Hatchability: None of the UV-B or UV-C exposed eggs in all treatments hatched while 90% control eggs hatched into viable larvae (Table 5). Eggs that failed to hatch showed little or no differentiation compared to controls when sub illuminated using light-level microscopy. Needham et al. (2006) exposed Dermatophagoides farinae eggs to UV-C for either 5 or 15 sec at less than 2 cm distance and noticed that the UV-C has potential to break the life cycle of this mite by killing the embryonic stage. Pulsed ultraviolet light at a wave length between approximately 200 and 400 nm is effective for control of insects and mites, not only over a wide a variety of species, but for a variety of developmental stages (Lagunas-Solar, 1997).
CONCLUSION
These findings suggest that UV-C irradiance at 260 nm is a more effective non-chemical measure than UV-B at 315 nm for reduction of the T. putrescentiae populations. In essence, the irradiation becomes lethal to the mite as a result of “resonance” effect on the colouring pigments, as well as the photothermic effect of rapid energy input. This method has the capability to replace the use of chemical acaricides, leaves no residue and can be implemented either in small scale with portable system or in a larger scale by integrating it to the existing technologies capable of handling agricultural commodities such as packaging materials. In addition, because of its physical nature, there is no possibility of mites building-up resistance through genetic changes transferred to new generations as it occurs with most chemical pesticides (Lagunas-Solar, 1997).
REFERENCES
- Franzolin, M.R., W. Gambale, R.G. Cuero and B. Correa, 1999. Interaction between toxigenic Aspergillus flavus Link and mites (Tyrophagus putrescentiae (Schrank)) on maize grains: Effects on fungal growth and aflatoxin production. J. Stored Prod. Res., 35: 215-224.
CrossRefDirect Link - Goodsell, D.S., 2001. The molecular perspective: Ultraviolet light and pyrimidine dimers. Oncologist, 6: 298-299.
CrossRefDirect Link - Hughes, A.M., 1976. The Mites of Stored Food and Houses. 2nd Edn., H.M. Stationery Office, London, ISBN-10: 0112409091, Pages: 400.
Direct Link - Jurkiewicz, B.A. and G.R. Buettner, 1994. Ultraviolet light-induced free radical formation in skin: An electron paramagnetic resonance study. Photochem. Photobiol., 59: 1-4.
CrossRef - Krantz, G.W., 1955. Some mites inJurious to farm-stored grain. J. Econ. Entomol., 48: 754-755.
Direct Link - Matsumoto, T., T. Hisano, M. Hamaguchi and T. Miike, 1996. Systemic anaphylaxis after eating storage-mite-contaminated food. Int. Arch. Allergy Immunol., 109: 197-200.
PubMed - Needham, G., C. Begg and S. Buchanan, 2006. Ultraviolet C exposure is fatal to American house dust mite eggs. J. Allergy Clin. Immunol., 117: S28-S28.
CrossRefDirect Link - Shindo, Y., E. Witt, D. Han and L. Packer, 1994. Dose-response effects of acute ultraviolet irradiation on antioxidants and molecular markers of oxidation in murine epidermis and dermis. J. Invest. Dermatol., 102: 470-475.
PubMed - Suzuki, T., M. Watanabe and M. Takeda, 2009. UV tolerance in the two-spotted spider mite, Tetranychus urticae. J. Insect Physiol., 55: 649-654.
CrossRefPubMedDirect Link - Van Bronswijk, J.E.M.H. and R.N. Sinha, 1973. Role of fungi in the survival of Dermatophagoides (Acarina: pyroglyphidae) in house-dust environment. Environ. Entomol., 2: 142-145.
Direct Link - Tan, D.X., L.C. Manchester, M.P. Terron, L.J. Flores and R.J. Reiter, 2007. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res., 42: 28-42.
CrossRefPubMedDirect Link