
L.B. Silva
Programa de Pos-Graduacao em Agronomia-Fitotecnia, Universidade Federal do Piaui no Campus Profa, Cinobelina Elvas, Bom Jesus-PI, Brazil
Z.F. Xavier
Programa de Pos-Graduacao em Agronomia-Fitotecnia, Universidade Federal do Piaui no Campus Profa, Cinobelina Elvas, Bom Jesus-PI, Brazil
C.B. Silva
Mestrado em Biologia Vegetal, Universidade Federal do Mato Grosso do Sul, Brazil
O. Faccenda
Titular da UEMS e pos-doutorando na Faculdade de Educacao da Unicamp, Brazil
A.C.S. Candido
Doutorado em Ciencias Farmaceuticas-Universidade Federal do Parana, Brazil
M.T.L.P. Peres
Associada 3 da Universidade Federal do Mato Grosso do Sul, Brazil
Journal of Entomology
Year: 2012 | Volume: 9 | Issue: 2 | Page No.: 98-106







ABSTRACT
Yield loss in different crops due to the attack of various classes of insect pests is a worldwide problem. Sucking type Dysdercus maurus causes damage to many crop species. Diverse plant species have been studied in order to identify the active ingredients responsible for their insecticidal activity against pests of economic importance. To verify the insecticidal activity of the stem bark and crude resin of Croton urucurana Baillon 1864 (Euphorbiaceae) against Dysdercus maurus Distant 1901 (Hemiptera: Pyrrhocoridae), the methanolic (EMeOH) extract, dichloromethane fraction (FDM), ethyl acetate fraction (FAE) and crude resin were applied topically with micro- applicator on the dorsal side of the mesothoracic region of third instar nymphs and the ventral side of adult insects and incorporated into an artificial diet. The average adult mortality of Dysdercus maurus treated topically with 0.5, 1.0 and 2.0% of the extracts mentioned above was significantly higher compared to controls. The FAE showed high insecticidal potential, having a higher mortality rate, extending the duration of the larval stages and resulting in malformed adults. Third instar nymphs fed with artificial seeds containing 1% FAE died three days after the start of the experiment, even before the molt to the next instar. For all treatments the death rate was greater for immature forms. These findings may provide a useful beginning for the development of biopesticides. Further studies will be carried out on mating, fecundity and proteolytic activity.
PDF Abstract XML References Citation
Received: July 29, 2011;
Accepted: October 14, 2011;
Published: December 12, 2011
How to cite this article
L.B. Silva, Z.F. Xavier, C.B. Silva, O. Faccenda, A.C.S. Candido and M.T.L.P. Peres, 2012. Insecticidal Effects of Croton urucurana Extracts and Crude Resin on Dysdercus maurus (Hemiptera: Pyrrocoridae). Journal of Entomology, 9: 98-106.
DOI: 10.3923/je.2012.98.106
URL: https://scialert.net/abstract/?doi=je.2012.98.106
DOI: 10.3923/je.2012.98.106
URL: https://scialert.net/abstract/?doi=je.2012.98.106
INTRODUCTION
Crop production is subject to a number of factors causing reduced productivity and loss. Among them, insect pests are a major factor, affecting about 30% of agricultural output. Application of insecticides is the control tactic most used to mitigate the severe losses caused by insects. The reasons for this, particularly in tropical and subtropical regions, include: good efficiency, low cost, ease of use and lack of viable alternatives for control in warmer regions (White and Leesch, 1995; Cooper and Dobson, 2007). However, the indiscriminate use of insecticides can cause reduction in populations of beneficial insects, resurgence and eruption of pests and loss of effectiveness due to the selection of resistant populations (Guedes and Pereira, 2008).
Given the increasing problems with resistance and impact on non-target organisms related to the use of synthetic insecticides, there is an urgent need for the development of safer alternatives, such as insecticidal proteins that interfere with the digestive process (Carlini and Grossi-de-Sa, 2002; Montesdeoca et al., 2005) and the use of substances from the secondary metabolism of plants with insecticidal activity (Omar et al., 2000; Simoes et al., 2002; Medina et al., 2003; Akhtar and Isman, 2004; Di Toto Blessing et al., 2010; Colom et al., 2007, 2008).
The Euphorbiaceae is one of the least studied plant families and deserves special mention for being one of the most extensive phanerogam families, composed of about 300 genera and 7,600 species (Cronquist, 1981). In this family, the genus Croton has economic importance due to its content of essential oils and various biologically active substances such as terpenoids, flavonoids and alkaloids (Peres et al., 1997, 1998; Suarez et al., 2003; Anazetti et al., 2004; Fischer et al., 2004). It is the second largest genus, with nearly 1,200 species distributed in tropical and subtropical regions (Webster, 1994). In Brazil there are approximately 300 registered Croton species growing in forests, fields and savannas throughout the country.
The species Croton cajucara Benth (Euphorbiaceae) is rich in diterpenes demonstrated to inhibit the growth of Heliothis virescens Fabr (Kubo et al., 1991). Croton linearis Jacq. contains a diterpene toxic to an important pest of sweet potato, Cylas formicarius elegantulus Summers (Alexander et al., 1991). Studies by Almeida et al. (1999) demonstrated that the ethanolic extract of Croton tiglium Willd. can cause mortality of 99% in Sitophilus zeamais Motsch. Similarly, the dichloromethane and ethyl acetate fractions of Croton urucurana caused approximately 65% mortality in larvae of Anagasta kuehniella, due to action of the phenolic compounds catechin and gallocatechin (Silva et al., 2009). Okokon and Nwafor (2009) reports that, the root extract of Croton zambesicus possess effects antimalarial and Robert et al. (2010) reports the anticoagulant effect of leaf extracts. Asare et al. (2011) reports that Croton membranaceus ingestion does not produce general acute toxicity. However, its creatinine kinase lowering ability could be explored. Some compounds isolated from the bark of Croton oblongifolius showed broad cytotoxicity on all cancer cell lines tested (Pudhom and Sommit, 2011). The crude resin (dragon’s blood) obtained from Croton urucurana has a potential antifungal effect that can be explored for therapeutic advantage as an alternative treatment for dermatophytosis or in conjunction with other antimycotics to allow the use of lower doses avoiding problems such as side effects and or resistance (Gurgel et al., 2005).
Today, it is known that some Croton species have strong insecticidal activity and in some cases are more toxic to insects than pyrethrum, a substance found in chrysanthemum flowers (Fazolin et al., 2005) that is already commercialized as an insecticide in many parts of the world. Croton is known for its toxicity to insects including some stored product insect pests (Alexander et al., 1991; Silva et al., 2009). Considering the potential use of Croton urucurana this study aimed to evaluate the insecticidal action of methanolic extract, semipurified fractions and crude resin of Croton urucurana (Baillon 1864) (Euphorbiaceae), against the cotton stainer bug Dysdercus maurus (Distant 1901) (Hemiptera: Pyrrhocoridae), insects economically important as crop pests.
MATERIALS AND METHODS
Botanical material, crude resin, methanolic extract and semipurified fractions: Stem bark and crude resin of C. urucurana were collected in the region of Dourados, Mato Grosso do Sul, Brazil. The species was identified by Claudio Conceição, Biology Department, Federal University of Mato Grosso do Sul-UFMS, Campo Grande-MS and a sample was deposited (No. 5009) at the CGMS Herbarium .
Stem bark, dried at room temperature and ground, was extracted with methanol. After solvent evaporation in a rotary evaporator, a solid brown-reddish material, hereafter referred to as methanol extract (EMeOH), was obtained. Afterwards, the material was percolated through a silica gel 60 (200 g) column, using hexane, dichloromethane and ethyl acetate successively. This procedure afforded three semipurified Fractions: Hexane (FH), dichloromethane (FDM) and ethyl acetate (FAE). Crude resin was collected directly from the trunk.
Insect bioassays: A colony of Dysdercus maurus was started with 30 pairs captured in dome traps in the ITA90 cotton area at the Agronomy Department experimental field station, Federal University of Mato Grosso do Sul in the city of Dourados, MS.
The insects were reared at a medium temperature of 23°C, 75% relative humidity, with a 16 h light-8 h dark cycle. D. maurus’ life cycle is about 30-40 days from eggs to adult insects. Each female lays up to 100-200 eggs over seeds. The eggs hatch after 5-8 days and the insects develop through five nymphal stages in about 20-25 days.
The insects were kept in transparent plastic pots, covered with screen material, with a 2 cm layer of autoclaved sand at the bottom. The insects were fed with cotton seeds and had free access to water, stored in plastic flasks, located inside the pots. The seeds were put inside the pots during the mating period and up to first instar. From the second instar on, the insects were transferred to a clean pot once a week and the seeds were placed over the screen material that covered the pots (Staniscuaski et al., 2005).
Bioassay I and II: A range of concentrations (0.5, 1 and 2%) of test solutions was prepared from stock extracts and applied topically with micro- applicator on the dorsal side of mesothoracic region of third instar nymphs and the ventral side of adult insects. Groups of 20 insects, each treated with 5 μL extracts and confined in a glass jar formed one replicate and 5 μL of the solvent applied on 20 insects served as the control. All treatments were replicated five times. Post treatment nymphal mortality, nymphal duration, nature of adults emerged and adult survival were monitored.
Bioassay III: Third instar nymphs were fed on artificial seeds following the methodology of Staniscuaski et al. (2005). The artificial seeds contained 1% of the test extracts and evaluation of survival was performed through the adult stage. All treatments were replicated three times. The duration of each developmental stage was checked to assess whether the test materials affected the development of insects.
Bioassay IV: Adults were fed on artificial seeds containing 1% of the test extracts and crud resin and survival was analyzed daily for a period of fifteen days. All treatments were replicated three times and adult survival was monitored.
Statistical analysis: The data obtained were analyzed by ANOVA or Kruskal-Wallis. A comparison of treatment means was performed using the Dunnet or Mann-Whitney test, p<0.05.
RESULTS
The effect of Croton urucurana methanolic extract (EMeOH), crude resin, dichloromethane (FDM) and ethyl acetate fraction (FAE) applied topically was assessed by determining the number of dead individuals seventy-two hours after application.
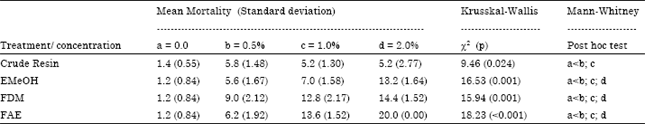
| Table 1: | Comparison of mean mortality in adult insects of Dysdercus maurus at 72 h after topical application of methanolic extract (EMeOH), dichloromethane (FDM), ethyl acetate fraction (FAE) and crude resin of Croton urucurana |
 | |
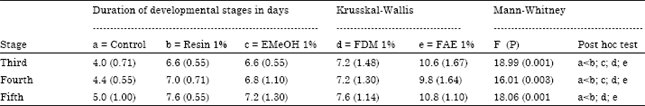
| Table 2: | Duration of developmental stages of Dysdercus maurus submitted to topical application of methanolic extract (EMeOH), dichloromethane fraction (FDM), ethyl acetate fraction (FAE) and crude resin of Croton urucurana, at concentrations of 1%. Results are expressed as means and standard deviations of the time required for molting of all the insects in each group |
 | |
The average adult mortality of Dysdercus maurus treated topically with 0.5, 1.0 and 2.0% of the compounds mentioned above was significantly higher than in the control group.
The mean percentage mortality of D. maurus increased with increase in concentration of the solutions tested, except for the crude resin. Topical application of the ethyl acetate fraction caused higher adult mortality (100%) compared with the other fractions (Table 1). The FAE fraction showed the strongest insecticidal activity.
D. maurus nymphs (third instar) were treated topically on the dorsal side of the mesothoracic region and the number of insects undergoing ecdysis in each group was counted for 15 days. All compounds significantly increased the average duration of all stages of development, except for fifth instar in the treatment with EMeOH (1%) (Table 2). The FAE fraction showed the highest insecticidal potential; it was associated with a higher mortality rate, increased duration of larval stages (Table 2) and a greater incidence of adult malformation.
In another experiment, third instar nymphs and adult insects were fed on artificial seeds containing C. urucurana methanolic extract (EMeOH), crude resin, dichloromethane (FDM) and ethyl acetate (FAE) (1%). It appears that the average rate of mortality was significantly higher for all treatments compared to control (Table 3). Third instar nymphs fed with artificial seed containing FAE 1% died three days after the start of the experiment, even before the molt to the next instar. For all treatments mortality was greater for immature forms. The adult insects, however, were susceptible to the same compounds as the immature insects (Table 3).
Figure 1 shows that the FAE fraction induced a more rapid entomotoxic effect in both third instar nymphs and adult insects, causing approximately 90% mortality in the first three days. The FDM fraction showed a mortality rate above 90% after the sixth day. The other treatments did not reach 90% mortality for immature forms.
Thus after a 5-10 day feeding period the mortality rate of insects fed on a diet containing 1% of C. urucurana methanolic extract (EMeOH), crude resin, dichloromethane (FDM) or ethyl acetate (FAE) was about threefold higher than that of the group feeding on a control diet.
| Table 3: | Effect on the rate of daily mortality of Dysdercus maurus, comparing third instar nymphs and adult insects fed on artificial seeds containing 1% methanolic extract (EMeOH), dichloromethane fraction (FDM), ethyl acetate fraction (FAE) or crude resin of Croton urucurana |
 | |
| Results are expressed as means and standard deviations of average mortality rate in third instar nymphs and adult insects in each group (* p<0.001) | |
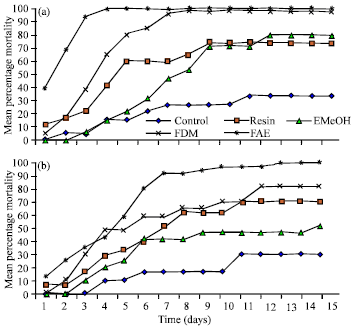 | |
| Fig. 1(a-b): | Insecticidal effect of methanolic extract (EMeOH), dichloromethane (FDM), ethyl acetate (FAE) fraction and crude resin of Croton urucurana on Dysdercus maurus. Nymphs (third instar) (a) or adults (b) fed on artificial seeds containing 1% of test substances\. The number of dead insects was counted daily. Results are expressed as means and standard deviations of five independent experiments, with N = 20 |
In insects fed with seeds containing FAE 1% the mortality rate was 90% after the sixth day (Fig. 1b). Adult insects fed on artificial seeds containing 1% methanolic extract (EMeOH), dichloromethane (FDM) fraction or crude resin of Croton urucurana were less susceptible to the insecticidal action of these compounds in comparison to the immature insects.
DISCUSSION
The utility of any botanical in plant protection against insect pests depends upon its toxicity to the target organism and/or its effects on development and reproduction or any other factor that leads to a reduction in its population. The primary aim of this work was to investigate the insecticidal activity of methanolic extract, crude resin and semipurified fraction of C. urucurana on the mortality and developmental of Dysdercus maurus. The selection of C. urucurana was based on popular information, its bactericidal activity, chemical studies (Peres et al., 1997, 1998) and reported insecticidal activity (Almeida et al., 1999; Torres et al., 2008; Silva et al., 2009).
Ventral topical application of semipurified FDM and FAE fractions caused significant mortality to adult insects of Dysdercus maurus, especially FAE 2% which was associated with a 100% mortality rate. This was similar to the results observed by Silva et al. (2009), who reported that the FAE 2% fraction caused 100% mortality in larvae of Anagasta kuehniella.
In topical application, the mean percentage mortality in D. maurus increased in a concentration dependent manner for all the substances tested. The methanolic extract (EMeOH), dichloromethane fraction (FDM), ethyl acetate fraction (FAE) and crude resin of C. urucurana were more toxic to D. maurus than to A. kuehniella.
All extracts, fractions and crude resin tested increased the average duration of the third, fourth and fifth instars. The FAE fraction was most active, possessing better insecticidal action both in topical application and when incorporated into artificial diet. The use of plants that affect insect larval development is advantageous because when the lifecycle is interrupted at this stage there is a reduction in the insect population during the period in which plant damage is being caused (Hernandez and Vendramim, 1996). Losses caused by pest attack during grain storage can reach up to 10% (Obeng-Ofori and Amiteye, 2005). In the experiment conducted with third instar nymphs adult malformation resulted, probably because the entomotoxic substance caused some disturbance in the metabolism of insect. It is necessary to investigate the possible mode of action and substance responsible for the insecticidal action, starting with the FAE fraction.
The FAE fraction when incorporated into artificial diet and fed to third instar nymphs of D. maurus causes 100% mortality before the third day. The insecticidal effect was observed for all material tested both in topical application and when incorporated into artificial diet of the third instar nymphs and adults. The average rate of mortality is higher for the third instar nymphs, probably due to difficulty degrading the insecticides. Studies revealed distinct proteolytic activities in the intestines of young and adult D. peruvianus (Staniscuaski et al., 2005; Piovesan et al., 2008). The difference in susceptibility to the compounds tested in third instar nymphs and adults of Dysdercus maurus may be due to decreased protein concentration in treated nymphs, as reported by Rao et al. (1999). To understand the internal physiological changes, the authors treated nymphs of Dysdercus koenigii with Artemisia annua oil and estimated the haemolymph protein concentration of fifth instar nymphs and adults emerged from them. It was observed that 3 days after treatment fifth instar nymphs had only 14.7 mg mL-1 protein concentration, compared with 24.9 mg mL-1 in control nymphs. This suggested that the treatment affected the protein level drastically during the early nymphal development. However, as development proceeded the difference in protein concentration between the treated and control insects narrowed. The treated fifth instar nymphs on day 6 after treatment showed a protein concentration of 20.8 mg mL-1 compared with 25.5 mg mL-1 in the control insects.
The adults that emerged from nymphs treated on days 3 and 6 had protein concentrations that were more or less equal to control insects, suggesting a restoration of protein level with increase in age. It appears that the early physiological effect of the treatment was on protein synthesis. Thus the toxic effect is most pronounced early in the insect’s development.
Larvae of Anagasta kuehniella when fed with artificial diet containing the FDM (2%) or FAE (1%) fractions showed lower efficiency of conversion of ingested and digested food which meant that in these diets a smaller quantity of the food (energy) was used for biomass production by the insect. This is possibly due to the fact that in those fractions an increased amount of metabolic energy was used for degradation of toxins. This can be evidenced when comparing the values of metabolic cost which were higher for larvae fed diets containing the FDM fraction. Approximate digestibility was also greater for larvae fed diets containing this fraction, indicating differential activity of the digestive enzymes (Silva et al., 2009).
In the methanolic extract of Croton urucurana Baillon (Euphorbiaceae) a number of known compounds, such as acetyl aleuritolic acid, stigmasterol, b-sitosterol, campesterol, b-sitosterol-O-glucoside, sonderianin, catechin and gallocatechin were isolated and identate (Peres et al., 1997, 1998) which played a role in insecticidal activity. According to Winkel-Shirley (2001), the condensed tannins catechin and gallocatechin are phenolic compounds responsible for the astringency of many fruits as well as defense against pests. They act by binding to insect digestive proteins and serve as an important mechanism for plant defense (Oliveira et al., 2003).
In bioassays using artificial seeds containing 1% concentrations of methanolic extract (EMeOH), dichloromethane fraction (FDM), ethyl acetate fraction (FAE) or crude resin of Croton urucurana, only the FAE fraction caused mortality above 90% after the sixth day. It can be inferred that the toxic compounds present in the diet are cumulative and that the insect is inefficient in the degradation. Comparing the results of all the fractions, extracts and crude resin, tested in experiments with topical application or when incorporated in the diet, it appears there is a concentration of entomotoxic compound in the FAE fraction that causes higher mortality in third instar nymphs and adult insects and increases the duration of the instars. Similar results were observed in the study of Silva et al. (2009).
Even with these favorable findings, additional toxicological studies with the tannin catechin should be made to assess its impact, especially on non-target species and to develop stable and convenient formulations of this potential phytoinsecticide (Silva et al., 2004). When insecticidal activity is detected, the active compounds are isolated, identified and subsequently used in large scale. In the laboratory, the new insecticide can be chemically transformed to suppress or minimize toxicity to mammals and natural enemies (when necessary). However, the choice of method is based on the complexity of the chemical structure of the substance and its synthesis may or may not be technologically or economically feasible (Fazolin et al., 2005).
The results presented in this report suggest that insecticidal extracts of C. urucurana could be an important component of IPM (integrated pest management), due not only to their ability to control survival rate and time of development but also by resulting in malformed adults. Further studies will be carried out on mating, fecundity and proteolytic activity.
REFERENCES
- Akhtar, Y. and M.B. Isman, 2004. Comparative growth inhibitory and antifeedant effects of plant extracts and pure allelochemicals on four phytophagous insect species. J. Applied Entomol., 128: 32-38.
CrossRefDirect Link - Colom, O.A., I. Barrachina, I.A. Mingol, M.C.G. Mas, P.M. Sanz, A. Neske and A. Bardon, 2008. Toxic effects of annonaceous acetogenins on
Oncopeltus fasciatus. J. Pest Sci., 81: 85-89.
CrossRef - Colom, O.A., A. Neske, S. Popich and A. Bardon, 2007. Toxic effects of annonaceous acetogenins from Annona cherimolia (Magnoliales: Annonaceae) on Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Pest Sci., 80: 63-67.
CrossRefDirect Link - Blessing, L.D.T., O.A. Colom, S. Popich, A. Neske and A. Bardon, 2010. Antifeedant and toxic effects of acetogenins from Annona montana on Spodoptera frugiperda. J. Pest Sci., 83: 307-310.
CrossRefDirect Link - Carlini, C.R. and M.F. Grossi-de-Sa, 2002. Plant toxic proteins with insecticidal properties. A review on their potentialities asbioinsecticides. Toxicon, 40: 1515-1539.
PubMed - Cooper, J. and H. Dobson, 2007. The benefits of pesticides to mankind and the environment. Crop Prot., 26: 1337-1348.
CrossRefDirect Link - Fischer, H., T.E. Machen, J.H. Widdicombe, T.J. Carlson, S.R. King, J.W. Chow and B. Illek, 2004. A novel extract SB-300 from the stem bark latex of Croton lechleri inhibits CFTR-mediated chloride secretion in human colonic epithelial cells. J. Ethnopharmacol., 93: 351-357.
PubMed - Asare, G.A., A. Sittie, K. Bugyei, B.A. Gyan and S. Adjei, 2011. Acute toxicity studies of Croton membranaceus root extract. J. Ethnopharmacol., 134: 938-943.
PubMed - Gurgel, L.A., J.J.C. Sidrim, D.T. Martins, V.C Filho and V.S. Rao, 2005. In vitro antifungal activity of dragon's blood from Croton urucurana against dermatophytes. J. Ethnopharmacol., 97: 409-412.
CrossRef - Kubo, I., Y. Asaka and K. Shibata, 1991. Insect growth inhibitory nor-diterpenes, cis-dehydrocrotonin and trans-dehydrocrotonin, from Croton cajucara. Phytochemistry, 30: 2545-2546.
CrossRefDirect Link - Medina, P., G. Smagghe, F. Budia, L. Tirry and E. Vinuela, 2003. Toxicity and absorption of azadirachtin, diflubenzuron, pyriproxyfen and tebufenozide after topical application in predatory larvae of Chrysoperla carnea (Neuroptera: Chrysopidae). Environ. Entomol., 32: 196-203.
Direct Link - Montesdeoca, M., M.G. Lobo, N. Casanas, A. Carnero, P. Castanera and F. Ortego, 2005. Partial characterization of the proteolytic enzymes in the gut of the banana weevil, Cosmopolites sordidus and effects of soybean Kunitz trypsin inhibitor on larval performance. Entomol. Exp. Appl., 116: 227-236.
CrossRef - Obeng-Ofori, D. and S. Amiteye, 2005. Efficacy of mixing vegetable oils mixed with pirimiphos-methyl against the maize weevil, Sitophilus zeamais Motschulsky in stored maize. J. Stored Prod. Res., 41: 56-57.
Direct Link - Okokon, J.E. and P.A. Nwafor, 2009. Antiplasmodial activity of root extract and fractions of Croton zambesicus. J. Ethnopharmacol., 121: 74-78.
CrossRefDirect Link - Oliveira, A.S., J. Xavier-Filho and M.P. Sales, 2003. Cysteine proteinases and cystatins. Braz. Arch. Biol. Technol., 46: 91-104.
CrossRef - Peres, M.T., F.D. Monache, A.B. Cruz, M.G. Pizzolatti and R.A. Yunes, 1997. Chemical composition and antimicrobial activity of Croton urucurana Baillon (Euphorbiaceae). J. Ethnopharmacol., 56: 223-226.
PubMed - Piovesan, A.R., F. Staniscuaski, J. Marco-Salvadori, R. Real-Guerra, M.S. Defferrari and C.R. Carlini, 2008. Stage-specific gut proteinases of the cotton stainer bug Dysdercus peruvianus: Role in the release of entomotoxic peptides from Canavalia ensiformis urease. Insect Biochem. Mol. Biol., 38: 1023-1032.
PubMed - Pudhom, K. and D. Sommit, 2011. Clerodane diterpenoids and a trisubstituted furan from Croton oblongifolius. Phytochem. Lett., 4: 147-150.
CrossRef - Robert, S., C. Baccelli, P. Devel, J.M. Dogne and J. Quetin-Leclercq, 2010. Effects of leaf extracts from Croton zambesicus Muell. Arg. on hemostasis. J. Ethnopharmacol., 128: 641-648.
CrossRefDirect Link - Winkel-Shirley, B., 2001. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology and biotechnology. Plant Physiol., 126: 485-493.
CrossRefDirect Link - Silva, H.H., I.G. Silva, R.M. dos Santos, E.R. Filho and C.N. Elias, 2004. Larvicidal activity of tannins isolated of Magonia pubescens St. Hil. (Sapindaceae) against Aedes aegypti (Diptera, Culicidae). Rev. Soc. Bras. Med. Trop., 37: 396-399.
PubMed - Silva, L.B., W. Silva, M.L.R. Macedo and M.T.L.P. Peres, 2009. Effects of Croton urucurana extracts and crude resin on Anagasta kuehniella (Lepidoptera: Pyralidae). Braz. Arch. Biol. Technol., 52: 653-664.
Direct Link - Torres, M., J. Assuncao, G. Santiago, M. Andrade-Neto and E. Silveira et al., 2008. Larvicidal and nematicidal activities of the leaf essential oil of Croton regelianus. Chem. Biodivers., 5: 2724-2728.
CrossRef - Webster, G.L., 1994. Synopsis of the genera and suprageneric taxa of Euphorbiaceae. Ann. Mo. Bot. Gard., 81: 33-144.
CrossRefDirect Link