
T. Ekrakene
Department of Basic Sciences, Faculty of Basic and Applied Sciences,Benson Idahosa University, P.M.B. 1100 Benin City, Nigeria
B. N. Iloba
Department of Animal and Environmental Biology, Faculty of Life Sciences,University of Benin, Benin City, Nigeria
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 1 | Page No.: 27-39







ABSTRACT
The decomposition processes of 48 white pigs (Sus scrofa Linnaeus) with mean weight of 22.3±1.20 kg (Mean±SD) resulting from four death agents (oxygen deprivation, slaughtering, aluminium phosphide and monocrotophos poisonings) was monitored throughout the Wet and Dry seasons of 2006/2007 and 2007/2008 seasons at Benson Idahosa University Demonstration farm, Benin City on Latitude 06°17'01.6"N and Longitude 05°36'10.6"E, 73 m of elevation from sea level. The insect fauna and periods of invasions were recorded. A consistent pattern of decomposition and insect invasion in both the dry and wet seasons regardless of killing agents was observed. The consistent insects from the earliest to the latest arrival being; Lucilia sericata; Chrysomya rufifacies, Musca domestica, Sarcophaga carnaria, Hermetia illucens and Ophyra aenescens from the order Diptera; Dermestis maculatus and Necrobia rufipes from the order Coleoptera. The other insect order, Hymenoptera was represented by members of the Formicidae (ants) family which was opportunistic. It also revealed that, the insects’ species that bred (Lucilia sericata, Chrysomya rufifacies, Musca domestica, Sarcophaga carnaria, Hermetia illucens, Ophyra aenescens and Dermestis maculatus) within the decomposition process in either seasons, all had one complete generation (egg, larva, pupa and adult or larva, pupa and adult) without an emerged adult from the process initiating and completing another cycle. This suggests that, the phenomenon of decomposition of animals is predictable from insect evidences since carrion insects seem to be decomposition-stage dependent, with only one generation of such insects’ species guaranteed, except where unpredictable circumstances present themselves. This understanding would enhance the global acceptability and applicability of testimonies from entomological data.
PDF Abstract XML References Citation
Received: January 26, 2010;
Accepted: April 23, 2010;
Published: July 17, 2010
How to cite this article
T. Ekrakene and B. N. Iloba, 2011. One Death, Many Insects’ Species Yet One Insect’s Generation. Journal of Entomology, 8: 27-39.
DOI: 10.3923/je.2011.27.39
URL: https://scialert.net/abstract/?doi=je.2011.27.39
DOI: 10.3923/je.2011.27.39
URL: https://scialert.net/abstract/?doi=je.2011.27.39
INTRODUCTION
The environment around man is predominantly a world dominated by arthropods and therefore, it is not strange to come in contact with arthropods as creatures. They contribute very immensely to the continuity of the earth, as they pollinate, eat other arthropods, both living plants and animals as well as their dead remains. Arthropods that feed on dead vertebrate bodies including man and other animals (carrion feeder) make them significantly important in forensic science. By their feeding action, they form part of the natural recycling of organic matter in the ecosystem in which they are found.
Man has dominated the ecosystem and modified the natural processes of the earth at will. According to Aggarwal (2005), nature has its own ways and one of its finest intricacies is the food web, that is, the process of eating and being eaten. He stated that the process is so immaculate that even dead bodies of animals, including those of humans, are decomposed by other creatures, with insects playing a predominant role. For centuries, until in the recent time in the developed world, Worms-maggots, either dead or crawling in the natural orifices (eyes, nose, anus, ears) and wounds on dead bodies were considered disgusting element of decay as soon as corpses were placed on table for autopsy. These maggots were most time washed away without any form of relevant information deduced from them. According to Catts and Haskell (1990), while ballistics, firearm examination, bitemarkes, gunpowder residue chemistry, blood splatter analysis and other elements of scientific criminology were studied and refined, the insects associated with death scenes were largely ignored. This however, has given a lot of drive to new frontiers in the field of Forensic Entomology which according to American Board of Forensic Entomology (1997) is the science of using insect evidence to uncover circumstances of interest to the law, often related to a crime.
Different researchers in carrion ecology have used different types of animals. Pigs are one of the most common animals used in this kind of research (Gill, 2005), but rabbits, cats, dogs, mice, birds, turtle and even elephant have also been used to study the decay process and insect succession on carrion (Payne, 1965; Denno and Cothran, 1976; Coe, 1978; Abell et al., 1982). With adequate knowledge of carrion insects which have shown regular successional pattern from literatures (Fuller, 1934; Payne, 1965; Braack, 1981; Early and Goff, 1986; Smith, 1986) insects associated with human corpses can represent a suitable Biological clock if the successional patterns are adequately developed for all geographic regions of the globe. Lefebvre (2009) has chattered a chronological succession pattern of necrophagous insects in human corpses as a new path in forensic entomology. This will in no doubt heighten the quest for standard path and procedures in the application of entomological data. The authenticity of entomological data is seriously being improved upon. This is obvious as researchers have started DNA based identification test. Notably, Wells et al. (2007) and Wells and Stevens (2008) have emphasized the importance of DNA based methods in forensic entomology. This according to them is to enhance identification of taxonomic groups validate insect evidence used in a death or other forensic entomological investigation. All these efforts are among others to enhance the general application and acceptability of insect evidence.
Based on available Forensic Entomology data, countries like the United States of America, United Kingdom, Australia and Germany have adopted insect evidences and so much sanity have been infused into their legal system as it concerns criminal tendencies that border around murder. Awareness is being created in countries like India and Pakistan where possible application of this new field of science is being considered. More investigators are becoming more and more aware of the invaluable contributions the silent testimony of insects can help to achieve in the legal system in bringing culpable offenders to book. According to Ekanem and Usua (1997) full-blown forensic entomology work did not start in Nigeria until 1988. Though, Usua (2007) noted that forensic entomology is at its infancy stage in Nigeria, he highlighted the possibilities of establishing the PMI of homicide cases with some certainty. In order for insect testimony to become global procedures with general acceptability and application, insects’ species generation obtainable from complete decomposition of carrions need to be addressed with the prevailing weather conditions. It is in this light therefore that, this study is aimed at determining the possible number of insect species generations that could occur at the prevailing local weather conditions in Benin City, Nigeria.
MATERIALS AND METHODS
Study site: The study site was the Demonstration Farm of Benson Idahosa University, Ugbor Village, Benin City, Edo State. The area lies within the rainforest vegetation belt of Nigeria and is characterized by two district seasons which run from November to March for wet and April to October for dry each year. The Latitude of the study area is 06°17'01.6"N and Longitude 005°36'10.6"E on an elevation of 73 m obtained from GPSmap76Csx Global Positioning System model.
Pig specimens used: Catts and Goff (1992) recommended 22 kg as the pig weight suitable for human model for adult decomposition. A total of 48 white pigs (Sus scrofa L.), were purchased from a piggery farm in Ekae village, Sapele Road, Benin City.
Killing exercise of pig specimens and experimental layout: Four killing methods were adapted from June 2006 to March 2008 with each method corresponding to each pig set.
| Set A: | Three Pigs killed by admixture of aluminum phosphide (food poisoning experiment). This killing method was used to simulate death occurring from poisoned foods and this pesticide being common among individuals for the killing of rats, a notable household pest |
| Set B: | Three Pigs killed by slaughtering method (cutlass used to cut neck region). This killing method was used to simulate death occasioned by cutting as this death method is common in rural settlements |
| Set C: | Three Pigs killed by injecting with 5 mL of monocrotophos poison (venous poisoning). This death type typified death from venous infusion of poison |
| Set D: | Three Pigs killed by oxygen deprivation (control experiment). This death method was used to simulate a natural cause of death |
Dead pigs were immediately packed in a separate heavy-duty polythene trash bag, labeled and transported to the study site. The times of death and deposition were noted on each daily data collection sheet corresponding to the pigs. The experimental layout was designed with distance between carrions being at least 15-20 m. Wire mesh of 20 mm mesh size and measuring 80 cm long x50 cm wide x60 cm high was placed over each pig to protect it from large vertebrate scavengers (Slone et al., 2005). The wire mesh used in the form of wire cages were secured with blocks around the carrions. These were removed and set aside at every sampling time.
Observations and daily data collections: Temperature and insect data collections were made at least once daily during the warm seasons. Observations and data collection were made 2 or 3 days interval on cooler season between the hours of 2-6 pm in the afternoon when most flies are active (Slone et al., 2005).
During the warm seasons however, sampling for insects were carried out in the morning (7-10 am) and during the afternoon (1-3 pm). This was to ensure that all ranges of insects were sampled as some flies maybe too active to be sampled during the noon period. Sweep net was used to collect flies while sampling for beetles and other crawling insects was done by carefully examining debris around the deposited pigs from 30-50 cm from the carcasses.
This exercise was constantly repeated until all fleshy tissues were exhausted from the carcasses. During these sampling periods, the pig specimens were not moved or disturbed, except for careful temperature readings that were taken from exposed maggot masses.
On deposition day after the pigs were positioned and weather data collected, collection of adult flies was made over each pig with side to side sweeping motions of a 12 inch sweep net. Flies collected were placed in vials with 70% alcohol and later pinned and identified.
In subsequent collections, sweep net was used similar to the first day. Eggs and larvae were sampled from the natural orifices such as ear, nostrils, eyes, anus, mouth and the cut region from the slaughtered carrions at different stages of decomposition. This was achieved by the careful examination of each using forceps to pick up the eggs and larvae into separately labeled rearing kits designed for the purpose. This was to enhance identification of implicated insects to species taxonomic level.
Half of the larvae collected were killed in boiled water before being placed in preservative solution to avoid shrinkage as demonstrated by Tantawi and Greenberg (1993). Thereafter, they were placed in vials with 70% alcohol for preservation. The third instar larvae were used for the identification to species taxonomic level. The remaining half of the sampled larvae in each sampling exercise, were placed in containers and reared to the adults stages, at the Biological Science Laboratory of Benson Idahosa University
Laboratory rearing of insects: Maggot rearing was carried out in the Biological Science Laboratory of Benson Idahosa University. Easton and Smith (1970), Nuorteva (1970) and Fisher (1980) had successfully used the laboratory method of insect rearing and found it easier for the purpose of identification.
Maggot rearing was achieved with plastic containers which were half-filled with substrate. The containers’ lids were mainly muslin cloth held tightly with rubber band. The choice of muslin cloth was to ensure that air circulated and the use of the rubber band helped to prevent larvae and adult flies from escaping through lids and also to prevent other flies entrance into the containers (Slone et al., 2005). The substrate was formulated from a mixture of dried leaves and branches of grasses and sandy soil. The plant mixture was ground thoroughly using an electric blender.
A few maggot samples from different orifice (ear, nostril, eye, cut region, mouth and anus) were placed on different rearing media, which were pieces of liver/tissue of pigs purchased from the market (Aggarwal (2005)). The containers were labeled according to pig sets and orifice from where the maggots were sampled and observations made several times to note changes in larval, pupation and emergence of adult insects. The containers of live maggots were monitored in the laboratory daily with prevailing daily temperature 28±2°C and 70% relative humidity.
RESULTS
Stages and decomposition time of pig carrions: The mean length of time in days taken by the carrions resulting from varied death agents at the wet (Fig. 1) and dry (Fig. 2) seasons are shown. During June 2006 to March 2008 seasons, a consistent pattern of decomposition was observed. Even though they showed a continuous process of decomposition, the process was easily divided into four distinct stages with unique significant insect groups and stages. The four decomposition stages evident were fresh decay stage, bloated, wet and dry decay stages. The observation showed that there were variations in the length of time spent in each of the stages.
For the wet season, fresh decay stage recorded least decomposition time and ranged from 2-4 days irrespective killing agents while dry stage recorded period of 35-42 days to be the longest process (Fig. 1). A similar pattern was equally observed for the dry season but showed general lower period of decomposition.
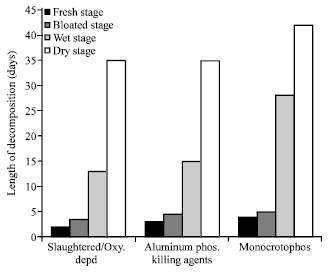 | |
| Fig. 1: | Average length of time in days taken by pig carcasses at the different decompositional stages during the wet seasons |
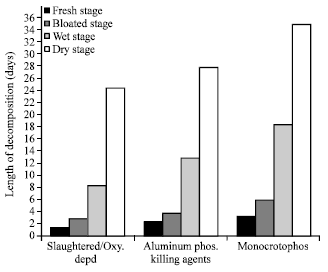 | |
| Fig. 2: | Average length of time in days taken by pig carcasses at the different decompositional stages at the dry seasons |
While the fresh decay stage recorded the shortest decomposition time ranging from 1.5-3.5 days, the dry decay stage recorded 24.5-35 days to be the longest decay stage independent of killing agents (Fig. 2).
In general, the dry season recorded averagely a shorter decomposition process in all the stages compared to the wet season. Irrespective of seasons (wet or dry) however, the dry decay stage of decomposition generally recorded the longest decompositional period (19-35 days) while the fresh decay stage was generally least (1-4 days). The bloated decay stage recorded mean decompositional period of 3-6 days and 9-28 days for the wet decay stage.
On the basis of killing agents, monocrotophos poisoned carrions recorded the longest decompositional time of 64-75 days independent of the season and was followed by sets of carrions killed from aluminum phosphide poison, 49-53 days. Slaughtered carrions and oxygen deprived carrions (control experiment) both recorded the least decomposition period of 32-45 days irrespective of the seasons (Fig. 1, 2).
| Table 1: | Summary of insects succession on carrions at seasons |
 | |
From the view point of seasons, the dry season recorded average shorter decomposition period compared to the wet season.
Waves of insects’ successional pattern on the pig carrions: Table 1 represents the insects’ successional pattern observed in this work and was found to be consistent and independent of the seasons. It revealed three distinct invasional patterns, which translated into the successional pattern of insect species from fresh decay to dry decay stages of carrion decomposition observed in this investigation. The first invasional event was that exploited by adult blowflies Lucilia sericata and Chrysomya rufifacies as well as adults of Musca domestica and Sarcophaga carnaria (Dipterans) which dominated from 1st-7th day of exposure of carrion depending on the agent of death. Within this period, the flies laid eggs and mated on the carrions. This first invasional wave of insect was succeeded by a second that was characterized by a large population of dipterans (L. sericata, C. rufifacies and Hermetia illuscens) larvae followed by invasion of adult beetles (Coleopterans). The presence of the Dipteran larvae also increased the invasion of ants (Hymenopterans) which were present, even at the first invasional event. At the second insects’ invasion, insect diversity and population was at the peak and degradation of carrion proceeded at a very fast rate. The insect diversities included all Dipterans (L. sericata, C. rufifacies, M. domestica, S. carnaria, H. illuscens and Ophyra aenescens) at different stages of development; Coleopterans (adults D. maculatus and N. rufipes) and ants. This second event lasted from 4th-30th day of exposure and was dependent on both weather conditions and killing agents. The second event of insect richness was proceeded by a sudden decline in population especially the larvae of Dipterans. This was as a result of the migratory activities of the pre-pupal stage and their subsequent development into adults. The third identified insect wave was dominated by Coleopterans and Hymenopterans (ants).
Daily mean temperature recorded for active decomposition with seasons: The main daily temperatures recorded for 30 days of active decomposition periods for the wet and dry seasons and the subsequent comparison with the Benin Weather Station (BWS) data are represented in Fig. 3. The field observations revealed that the dry seasons showed general higher daily temperatures which ranged from 34.3 to 37.1°C compared to the wet seasons which recorded 23 to 31.3°C. Also, while the dry season recorded a mean value of 36.1°C, the wet season had a mean value of 27.2°C.
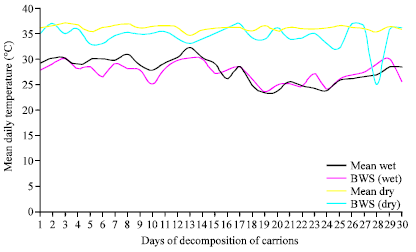 | |
| Fig. 3: | Mean daily temperature (°C) recorded for 30 days of active decomposition in the Dry (November) and Wet (June) seasons compared with Benin Weather Station (BWS) |
DISCUSSION
This investigation revealed descriptive decomposition stages. Though, Schoenly and Reid (1987) defined decomposition as a continuous process without discrete stages, many other workers have divided the process into four to six stages (Payne, 1965; Lord and Burger, 1984; Morris, 1988; Tantawi et al., 1996). In this present study, the decomposition process was indeed continuous but showed remarkable evidence in four distinct stages of decay which are successive. These successive and distinct stages were fresh, bloated, wet decay as well as dry decay. These stages were distinctly marked by characteristic odour, presence or absence of insect, the stage and type of insect species present as well as the insect activity syncronising with the insect present. These observations agreed in many respect to the findings of earlier researchers, notably, Reed (1958), Rodriguez and Bass (1983), Morris (1988) and Tantawi et al. (1996) as well as Parikh (1999), Vij (2001) and Pillay (2004).
Array of insects from different orders have been associated with the decomposition processes of carrions. Smith, 1986 categorized insects associated with carrion on the basis of their food preferences and their ecological role. Gill (2005) emphasized that, these insects species typically occur in succession and respond to decompositional changes of the carcass. This present study recorded insects species from three insect orders; Diptera, Coleoptera and Hymenoptera. The occurrence and succession of Lucilia sericata, Chrysomya rufifacies, Musca domestica, Ophyra aenescens, Hermetia illuscens, Sarcophaga carnaria (Diptera); Dermestis maculatus and Necrobia rufipes (Coleoptera) and the opportunistic ants (Hymenoptera) were found to be independent of both season (wet and dry) and the killing agents. The findings in this present work, that all the implicated insects were from Diptera, Coleoptera and Hymenoptera concurred with most earlier investigators including Payne (1965), Smith, 1986, Keh (1985), Lord (1990), Catts and Goff (1992), Anderson (2000) and Aggarwal (2005) and others.
The families of the Diptera order represented in this study include; Calliphoridae, Muscidae, Sarcophagidae and Stratiomydae while the Coleoptera order was represented by the Cleridae and Dermestidae families. The order Hymenoptera was represented by the ants (Formicidae). These families were seen to be consistent in their occurrence and have a regular sequence of occurrence. The families of insects implicated in this study have been highlighted by other researchers to be involved in decomposition processes of carrions.
The two species of Calliphoridae represented in this present study Lucilia sericata and Chrysomya rufifacies were among the earliest Dipterans that visited the pig carcasses. These two species were also the only ones that formed maggot masses, thus, resulted in the fast degradation of the carrions. These two species were the predominantly sampled insect in both seasons and in all carrions resulting from the two killing methods. Similar observations were made by Reed (1958), Johnson (1975), Tantawi et al. (1996), Bharti and Singh (2003) and Aggarwal (2005). Although Coe (1978) and Braack (1981) had noted that Chrysomyinae flies can act as primary species in the absence of Calliphorinae only; Flynn and Moorhouse (1979) and Bharti and Singh (2003) reported that, species from both sub-families can act as primary species whenever they breed on corpses. In this present study however, though both sub-families were represented by Lucilia sericata and Chrysomya rufifacies, L. sericata showed more dominance in number compared to C. rufifacies.
Chrysomya rufifacies according to Greenberg and Povolny (1971) is more adapted to tropical conditions and is found all year round. In the course of this investigation, similar observation was made and was also present in all carrions subjected to different killing agents. Many scientist including Goff et al. (1986), Goff and Odum (1987) as well as Singh and Bharti (2000) have used this insect species as a forensic indicator. The Muscidae family is cosmopolitan but species occurrence varies from region to region. In this present study, Muscidae was represented by Musca domestica and Ophyra aenescens. M. domestica arrived within 15 min after death and adult emerged from sampled larvae bred in the laboratory between 12 and 18 days. Though there was no maggot mass formed, it was observed to have bred on the pig carcasses. This is however in contrast to the observations made by Aggarwal (2005) that, though M. domestica, the common house fly was collected from corpses, it was not found to breed on them. Ophyra aenescens is originally an old world species, known to have a wide distribution in the world (Costa et al., 2000). In this present investigation, O. aenescens was consistent in the insect successional profile on the carrions irrespective of season and method. Their arrival time was consistently observed to be between 3-5 days and correspondent to the bloated decay stage of carrion decomposition. This agreed with the findings of Carrera (1991), who observed that muscids of the genus Ophyra are attracted by fermenting corpses with amoniacal emanations. Costa et al. (2000) noted that Ophyra species breed in numerous media and are known to invade corpses after other flies, such as Muscina (Muscidae) and Calliphora (Calliphoridae), but before the phorids (Phoridae). Smith, 1986 and Bourel et al. (2004) observed that Ophyra species are sometimes found on human bodies kept indoors for several months, where blowflies (Diptera, Calliphoridae) do not have access to them. Though they were present and had their complete development stages from egg to adult on the carrions which lasted between 21 to 28 days, their forensic role could not be ascertained in this investigation. However, Ophyra species have been reported in literature from buried corpses (Mégnin 1894 in Smith, 1986) and other forensic evaluations of crime histories with monospecific populations (Turchetto and Vanin, 2004), as Hydrotaea capensis (Bourel et al., 2004; Grassberger and Scharrer-Liška, 2006). Skidmore (1985) mentioned that some Ophyra species, belonging to the sub-family Azeliinae, are facultative or obligatory carnivores in the larval phase. Costa et al. (2000) also noted that members of this genus are frequently used in the biological control of M. domestica.
The family Stratiomyidae was represented by the Black Soldier Fly, Hermetia illucens. This insect was sampled within the first 7 days after death at the early wet decay stage. Its larvae are characteristically dorso-ventrally flattened and observed to churn up the abdominal fleshy remains in a characteristic manner. The precise oviposition time could not be ascertained from this investigation, but larvae bred from the laboratory indicated that emergence of adult occurred between 45 to 55 days. This was correspondent to the estimated period on carrion. It was however observed, that emergence was faster during the dry season where it took between 45 to 50 days while the wet season recorded 45 to 55 days. This insect species has been implicated as a forensic indicator by a few researchers. Lord et al. (1993) had reported that H. illucens initiates oviposition 20 to 30 days after death and its life cycle would require at least 55 days depending on temperature for completion. They concluded that H. illucens could provide valuable parameters to forensic investigators for estimating the PMI of badly decomposed remains. Also the information provided by the development time of H. illucens in combination with the time required by the ant Anoplolepsis longipes to establish a colony was used by Goff and Win (1997) to solve a case in which human remains were found inside a metal tool box.
The family Dermestidae was represented by Dermestis maculatus. These beetles are well known to feed on dry skin and bones (Payne and King, 1970). Putman (1977) considered them as true carrion feeders, playing important role in carcass degradation. Catts and Goff (1992) and Goff (1992) in their respective investigations on forensic entomology in criminal cases and estimation of post mortem interval using arthropod development and successional patterns have stated that Dermestis maculatus (hide beetles) have the potential to offer investigators an estimation of the time since death in homicide or questionable cases. They also observed that similar to the use of flies in forensic entomology, the arrival of D. maculatus to carcasses occurs in a predictable succession. Richards and Goff (1997) reported that adult D. maculatus beetles generally arrive 5 to 11 days after death. This ascertion by Richards and Goff (1997) is highly support by this present work as its arrival was observed between 5-10 days after death. It is one of the most abundant insects at the dry decay stage of decomposition, when the adults and larvae are involved in the consumption of skins regions left from previous decay stages. Payne and King (1970) have noted that larvae and adults associated with carrion feed on dried skin, sinews and bones. This insect has been reported in previous studies and has also been used as a forensic indicator (Benecke, 1998; Greenberg and Wells, 1998; Bharti and Singh (2003)). A significant observation made in the course of this present investigation was that, breeding of the D. maculatus was only observable during the dry seasons.
Necrobia rufipes, belonging to Cleridae family preceded D. maculatus in this study. Necrobia rufipes was observed during the early wet decay stage of decomposition. They were predominantly found in regions of carcasses that have achieved high level of dryness. Though, this species could not be found breeding on the carrion during this study, estimated period on carrion was not less than 3 weeks. The rains during the wet season reduced the time period the species spent on the carcasses as live adults were destroyed by the rains during this season. Also, the rain affected the sinews and skin by washing them away from the bones which hitherto acted as food for this species. This genus is the most reported by majority of the earlier workers from carrion (Aggarwal (2005)). Many researchers have used it as a forensic indicator (Goff and Odum, 1987; Lord, 1990; Bharti and Singh (2003)).
Members of the formicidae (ants) family were observed throughout this present study. Their occurrence was observed to be irrespective of seasons, killing method and stage of decomposition. Field observation revealed that their presence increased with increase in decomposition stages and insects development. They use carcass as food and also acted as predator to maggots, pupae and puparia cases. This may place them as adventives or incidental species. This suggests that, their presence is opportunistic and therefore unimportant in the decomposition of carrion. However, their predatory actions on insect, especially the eggs and the maggots have the potential of retarding the rate of decomposition. Similar observations have been made by Fuller (1934), Early and Goff (1986), Singh and Bharti (2000) and Aggarwal (2005).
The most critical and perhaps outstanding field observation from this research was the fact that, none of the implicated insects’ species had more than one generation on the decomposing carrions. They (insects species) all appeared at a different specific stage of the decomposition process (Table 1) and commenced the relevant stage of life, either by laying of eggs as in Lucilia sericata, Chrysomya rufifacies, Musca domestica, Hermetia illuscens and Dermestis maculatus or deposited first instar larvae as in Sarcophaga carnaria. Others such as Necrobia rufipes and the ants merely used the carrions as food resource without necessarily undergoing any form of development within the process. These developmental stages of the insects were consistently observed irrespective of seasons and were decomposition-stage dependent. From field observation, insects species involved in carrion decomposition seem to be synchronized with insect type and stages of development, it therefore followed that, an emerged adult from the decomposition process may not be able to successfully complete another cycle, should it have an opportunity of initiating one. The only way perhaps where another cycle could be completed may be in either buried carrions or carrions subjected to circumstances in which general decomposition is prolonged without affecting the carrion insects. This may need to be investigated for further insight. The simple deduction that could be obtained from this is that, insect species that visit carrions are ‘decomposition-stage dependent’ and once a given decomposition stage was over; the insects species specific to it would be unable to adequately adapt. This could mean that, the reproductive, physiologic and nutritional requirements that the insects need to be attracted to the decomposing remains of animals are not in place. This observation therefore has prompted the conclusion that “one death, many insects’ species, yet one generation” is a predictable routine except unpredictable circumstances interface. This reasoning seems vital and true and will enhance entomological evidences if properly harnessed. It will also enhance global uniformity of entomological data and interpretations except where unpredictable circumstances or conditions present themselves.
ACKNOWLEDGMENT
The management of Benson Idahosa University Benin City, Nigeria is acknowledged for the Ph.D. grant this work enjoyed.
REFERENCES
- Abell, D.H., S.S. Wasti and G.C. Hartmann, 1982. Saprophagous arthropod fauna associated with turtle carrion. Applied Entomol. Zool., 17: 301-307.
Direct Link - Anderson, G.S., 2000. Minimum and maximum development rates of some forensically important calliphoridae (Diptera). J. Forensic Sci., 45: 824-832.
PubMedDirect Link - Benecke, M., 1998. Random amplified polymorphic DNA typing of necrophagous insects (diptera, coleoptera) in criminal forensic studies: Validation and use in practice. Forensic Sci. Int., 98: 157-168.
Direct Link - Bharti, M. and D. Singh, 2003. Insect faunal succession on decaying rabbit carcasses in Punjab. J. Forensic Sci., 48: 1133-1143.
Direct Link - Bourel, B., G. Tournel, V. Hedouin and D. Gosset, 2004. Entomofauna of buried bodies in northern France. Int. J. Legal Med., 118: 215-220.
CrossRefDirect Link - Catts, E.P., 1992. Problems in estimating the postmortem interval in death investigations. J. Agric. Entomol., 9: 245-255.
Direct Link - Catts, E.P. and M.L. Goff, 1992. Forensic entomology in criminal investigations. Annu. Rev. Entomol., 37: 253-272.
CrossRefDirect Link - Costa, P.R.P., R.L. Franz, E.E.S. Vianna and P.B. Ribeiro, 2000. Synanthropy of Ophyra spp. (Diptera: Muscidae) in Pelotas state of Brazil. Brazil. J. Vet. Parasitol., 9: 165-168.
Direct Link - Denno, R.F. and W.R. Cothran, 1976. Competitive interactions and ecological strategies of sarcophagid and calliphorid flies inhabiting rabbit carrion. Entomol. Soc. Am. Ann., 69: 109-113.
CrossRefDirect Link - Early, M. and M.L. Goff, 1986. Arthropod succession patterns in exposed carrion on the island of O'Ahu, Hawaiian Islands, USA. J. Med. Entomol., 23: 520-523.
CrossRefDirect Link - Easton, A.M. and K.G.V. Smith, 1970. The entomology of a cadaver. Med. Sci. Law, 10: 208-215.
PubMedDirect Link - Goff, M.L., 1992. Problems in estimation of postmortem interval resulting from wrapping of the corpse: A case study from Hawaii. J. Agric. Entomol., 9: 237-243.
Direct Link - Goff, M.L., M. Early, C.B. Odum and K. Tullis, 1986. A preliminary checklist of arthropods associated with exposed carrion in the Hawaiian Islands. Proc. Hawaiian Entomol. Soc., 26: 53-57.
Direct Link - Goff, M.L. and C.B. Odum, 1987. Forensic entomology in the Hawaiian Islands: Three case studies. Am. J. Forensic Med. Pathol., 8: 45-50.
PubMedDirect Link - Goff, M.L. and B.H. Win, 1997. Estimation of post mortem interval based on colony development time for Anoplolepsis longipes (Hymenoptera: Formicidae). J. Forensic Sci., 42: 1176-1179.
PubMedDirect Link - Johnson, M.D., 1975. Seasonal and microseral variations in the insect populations on carrion. Am. Midland Naturalist, 93: 79-90.
CrossRefDirect Link - Keh, B., 1985. Scope and application of forensic entomology. Annu. Rev. Entomol., 30: 137-154.
Direct Link - Lefebvre, G., 2009. Forensic entomology: A new hypothesis for the chronological succession patterns of necrophagous insect in human corpses. Ann. Soc. Entomol., 45: 377-392.
Direct Link - Payne, J.A., 1965. A summer carrion study of the baby pig Sus scrofa Linnaeus. Ecology, 46: 592-602.
CrossRefDirect Link - Payne, J.A., E.W. King and G. Beinhart, 1968. Coleoptera associated with pig carrion. Entomol. Monthly Magaz., 105: 224-232.
Direct Link - Putman, R.J., 1977. Dynamics of the blowfly Calliphora erythrocephala within carrion. J. Anim. Ecol., 46: 853-866.
Direct Link - Reed, Jr.H.B., 1958. A study of dog carcass communities in tennessee, with special reference to the insects. Am. Midland Nat., 59: 213-245.
CrossRefDirect Link - Richards, E.N. and M.L. Goff, 1997. Arthropod succession on exposed carrion in three contrasting tropical habitats on Hawaii Islands, Hawaii. J. Med. Entomol., 34: 328-339.
PubMedDirect Link - Rodriguez, W.C. and W.M. Bass, 1983. Insect activity and its relationship to decay rates of human cadavers in east tennessee. J. Forensic Sci., 28: 423-432.
Direct Link - Schoenly, K. and W. Reid, 1987. Dynamics of heterotrophic succession in carrion arthropod assemblages: Discrete seres or a continuum of change? Oecologia, 73: 192-202.
CrossRefDirect Link - Smith, K.G.V., 1986. A Manual of Forensic Entomology. 1st Edn., British Museum (Natural History), London and Cornell University Press, Ithaca, Pages: 205.
Direct Link - Tantawi, T.I. and B. Greenberg, 1993. The effect of killing and preservative solutions on estimates of maggot age in forensic cases. J. Forensic Sci., 38: 702-707.
PubMedDirect Link - Tantawi, T.I., E.M. El-Kady, B. Greenberg and H.A. El-Ghaffar, 1996. Arthropod succession on exposed rabbit carrion in Alexandria, Egypt. J. Med. Entomol., 33: 566-580.
Direct Link - Turchetto, M. and S. Vanin, 2004. Forensic evaluations on the crime case with monospecific necrophagous fly population infected by two parasitoid species. Aggrawal`s Internet J. Forensic Med. Toxicol., 5: 12-18.
Direct Link - Wells, J.D., R. Wall and J.R. Stevens, 2007. Phylogenetic analysis of forensically important Lucilia flies based on cytochrome oxidase 1 sequence: A cautionary tale for forensic species determination. Int. J. Legal Med., 121: 229-233.
CrossRefDirect Link