
Hanan A. El-Sadawy
Department of Parasitology and Animal Diseases National Research Center, Cairo, Egypt
Amira El-Shazly
Department of Zoology, Faculty of Science, Cairo University, Cairo, Egypt
Rabab M.M. El-Khateeb
Department of Zoology, Faculty of Science, Cairo University, Cairo, Egypt
Journal of Entomology
Year: 2008 | Volume: 5 | Issue: 2 | Page No.: 62-76







ABSTRACT
Histopathological studies revealed a highly damaged midgut epithelial cells with different species of Heterorhabditids. Also, the presence of differentiated cells in all of the midgut regions, as seen from serial sections, suggests that digestive processes appear to occur throughout the length and diameter of the midgut and its diverticulae. The ultrastructural study of the gut showed that the granules of S1 and S2 secretory cells were different and the two cell types could sometimes be found together in one caecum.
PDF Abstract XML References Citation
How to cite this article
Hanan A. El-Sadawy, Amira El-Shazly and Rabab M.M. El-Khateeb, 2008. Histopathological Studies of Hard Tickes Hyalomma dromedarii Infected of Entomopathogenic Nematodes. Journal of Entomology, 5: 62-76.
DOI: 10.3923/je.2008.62.76
URL: https://scialert.net/abstract/?doi=je.2008.62.76
DOI: 10.3923/je.2008.62.76
URL: https://scialert.net/abstract/?doi=je.2008.62.76
INTRODUCTION
Studies during the past decade have shown that entomopathogenic nematodes are also pathogenic to ticks (Samish and Glazer, 1991). Of 16 ixodid tick species from six genera and three argasid species from the two genera tested, only one species was not susceptible to nematodes (Samish et al., 2000). Unfortunately, few literature concerning the effect of entomopathogenic nematodes on the histopathology of ticks were found. Light and electron microscopical analysis were used to detect Borrelia japonica and Boorrelia burgdrorferii in the tissue of ticks (Szabo and Bechara, 1999; Amosova, 2000). Another microscopical analysis studied the morphological changes of midgut which accompanied feeding and digestion of the blood meal in the midgut epithelium of B. microplus and Rhipicephalus appendiculatus (Walker and Fletcher, 1987; Agyei and Runham, 1995). Investigation of 1789 adult ticks in Egypt for the presence of microorganisms revealed the presence of protozoan diseases agent; Theileria annulata, Babesia bigemmina and the Rickettesia aegyptianella pullorum which were found in saliva and midgut of eight ixodid and two argasid tick species (El Kammah et al., 2004). This study aims to study the hisopathological changes resulting to entomopathogenic nematode infections. This study may be will answer a question, why ticks die although nematode does not complete its development inside it?
MATERIALS AND METHODS
Histopathological study on female ticks (H. dromedarii) exposed to entomopathogenic nematodes.
Light Microscopical Analysis
The engorged female ticks were exposed to five species of entomopathogenic nematodes: S. carpacapsae DD136, S. riobravae, H. bacteriophora HP88, H. indicus RM1 and H. bacteriophora Eg1. The ticks were placed individually in plastic pots containing moistened sand RH 20% (w.w). Ticks were infected with 4000 IJs/tick. The infection technique was carried out as previously described by Kocan et al. (1998).
Three ticks were collected at each 2, 4, 6 and 8 days post-infection and in fixed in 10% formalin. The staining method was carried out according to Agyei and Runham (1995). Sections of ticks at each interval were stained for general morphology with haematoxyline and eosin (H and E).
Ultrastructural Examination
Five female ticks of 6 mm length and 12 fully engorged female H. dromedarii ticks were collected. The 6 mm female ticks were dissected in Karnovsky’s fixative (2% glutaraldehyde, 2.5% paraformaldehyde, 0.1M phosphate buffer, 0.15M NaCl and 0.1 mM CaCl2, pH 7.2) at 4°C. Pieces of gut were transferred to fresh fixative and fixed at 4°C for 16 h. The engorged ticks were incubated at 28±1°C and >90% relative humidity for up to 4 days post engorgement. Three of these ticks were taken each day, dissected in the same fixative and small pieces of gut were transferred to fresh fixative for 16 h at 4°C. The gut pieces were then washed for 24 h in the phosphate buffer with the same NaCl and CaCl2 concentrations and post-fixed in 1% osmium tetroxide in the same buffer. Some of the tissues were stained en bloc with 1% uranyl acetate. Tissues were dehydrated in ethanol, embedded in eon-araldyte. Ultrathin sections were stained with uranly acetate and lead cirtrate (Reynolds’ methods Glauert, 1975) and observed in a Philips M300 electron microscope. Light microscopy of the gut cells was performed as described by Agbede and Kemp (1985).
RESULTS
Histopathological Study on Engorged Female Tick (H. dromedarii)
The Light Microscop
The morphological and histological characteristics of the midgut digest cells of H. dromedarii were in general similar to those described for B. microplus and R. appendiculatus. However, differences were found in the shape of the midgut epithelial cells especially in the prodigest and sessile digest cells. There was no pronounced differences in the progress of digestion in the different gut caeca with those described in Table 1. The development of cells from undifferentiated stem cells to a number of secretory and digestive cell types follows the nomenclature of as shown in Table 1.
Midgut Epithelial Cells Developmental
Changes on Feeding
On attachment and feeding, the midgut epithelium underwent dramatic changes characterized by loss of hematin and proliferation of the cells. The classification of the cells of the midgut follows the nomenclature of other researchers (Agyei et al., 1991; Agyei and Runham, 1995).
| Table 1: | Cell types in the gut of Ixodid ticks |
 | |
The cell types described by Hughes (1954), Till (1961), Chinery (1964) and Tatchel (1964) were discussed by Balashov (1972); +dhl: mature digest cells with haemoglobin and leucocytes; Dhv: mature digest cells with haemoglobin and no leucocytes | |
Stem Cell
This cell is usually dome- shaped and is found in the early stages of each instar, although it is most numerous at the mid-stage of nymphal and adult development. The cell is undifferentiated and cytoplasmic organelles cannot be distinguished by light microscopy. It was located on the basement membrane (Fig. 1 and 2).
Digest Cell
This was the dominant cell type in the midgut epithelium and seemed to originate directly from the stem cell (Fig. 1 and 2). It appeared as a cuboidal cell and the stages manifest during its development were: prodigest; sessile; residual sessile, empty; detached and spent digest cell phases. This nomenclature a part from prodigest and empty digest cells, have been adopted from Walker and Fletcher (1987). It is in the main descriptive and is based on observations from light microscopy supported with ultrastractural details.
Prodigest Cell
This was slightly elongate with round nucleus and a large deeply staining nucleolus (Fig. 2). The cytoplasm was intensely basophilic due to RNA accumulation which was lost with ribonuclease digestion as suggested by Agyei and Runham (1995). The basal membrane showed massive infoldings which were closely associated with large number of mitochondria. The cell appeared to absorb the blood meal hemoglobin and with this absorption, the prodigest cell transform into the sessile cell.
Sessile Digest Cell
This was comparatively large cell, also columnar in shape and it appeared soon after the intake of the first blood meal. The cell had a broad base in contact with the basement membrane in contrast with the cell described under the same name by Walker and Fletcher (1987) in R. appendiculatus.
Residual Sessile Digest Cell
This was generally more club-shaped though many shapes were seen and it probably developed from the sessile digest cell. Its name derives from the fact that while the sessile digest cells had their basal part in contact with the basement membrane, few cells, the residual cells, remained thinly attached to the basement membrane (Fig. 2) and appeared to move into the lumen.
Spent Digest Cell
This was the last of the digest cell series. It probably represented the last phase of digestive activity and in some cases was followed by autolysis. This resulted in the release of hematin and other indigestible products of intracellular digestion into the lumen (Fig. 2).
Empty Digest Cell
This was a quiescent digest cell type found most commonly in the molting and post-moulting phases of larvae and nymph and it may transform into the prodigest or sessile digest cell. It was hemispherical, with a moderate round nucleus and blackish-brown or violet colored hematin granules distributed throughout the cytoplasm.
Secretory Cell
The presence of a specific secretory cell has been reported in B. microplus (Agbede and Kemp, 1985) and R. appendiculatus (Agyei and Runham, 1995). However, in the present study, it was difficult to distinguish specific secretory cells morphologically from other midgut digest cells.
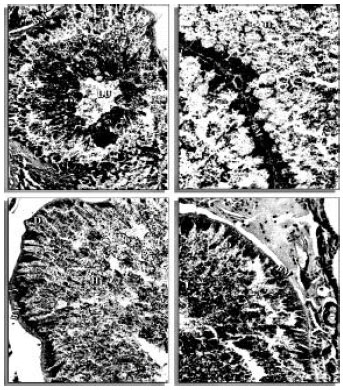 | |
| Fig. 1: | Transverse sections of the midgut of H. dromedarii showing lumen (LU), basement embrance (BM), digest cell (D) with release of hematin (H) by the midgut epithelial cells. (160x) |
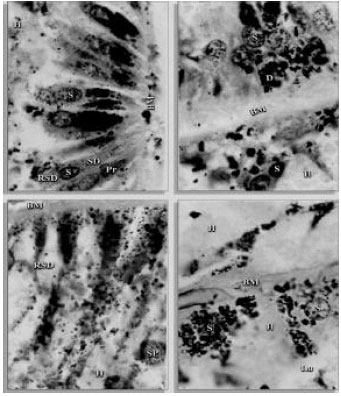 | |
| Fig. 2: | Transverse sections of the midgut of H. dromedarii showing basement membrance (BM), digest cell (D), predigest cell (Pr), sessile digest cell (SD), residual sessile digest cell (RSD), spent digest cell (SP), secretory cell (S), Lumen (Lu), hematin (H) (1000x) |
Gut Cell Development During Feeding of Females after Exposure with Different Nematode Species
Heterorhabditis Indicus RM1
A damage of the midgut epithelial cells appear after 2 days of infection. The gut was lumen surrounded by a thin unicellular layer of cuboidal digest cells which remain from the late stage. The digest and the residual sessile digest cells was not formed and the basement membrane was not damaged until the 2nd post-infection. Indeed, basement membrane was completely damaged after 4th and 6th days post-infection. The digest cell was the main cell type present and now the cell has various sizes and shapes. Many have enlarging nuclei, corresponding to the enlarging intermediate cells of the digest cell series. Moreover, no hematin in the digest cells and the midgut lumen filled with hemolysis blood could be observed. In response, cell types after 6th days post infection was hardly differentiated (Fig. 3A-D). Secretory cells were rarely seen within the epithelial layer and there were fewer digest cells.
Heterorhabditis bacteriophora HP88
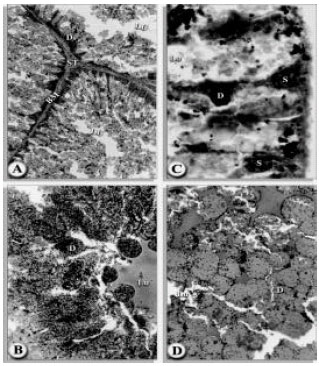
Less damage appeared in the midgut epithelial cell during the 2nd day post-infection with H. bacteriophora HP88 which increased significantly during the 4th day. The caeca showed stricking changes, where the cells were larger and possessed large nuclei. Stem cells enlarged to become digest cells. Maximal damage reached during the 6 days post-infection. A few of the digest cells were cuboidal in shape, while others were dome shaped and contain small food vacuoles which were fairly evenly distributed in the cells. Many of the cells have a few large food vacuoles and a few small dark brown hematin granules (Fig. 4A-D). Secretory cells were virtually absent. Stem cells were few but since some are in division there appears to be a new generation of stem cell forming.
Heterorhabditis bacteriophora Eg1:
A slight damage of the midgut cells appeared after two days post infection. The digest cells were all packed with food vacuoles, most of which are large and stain heavily with eosin. Some cells
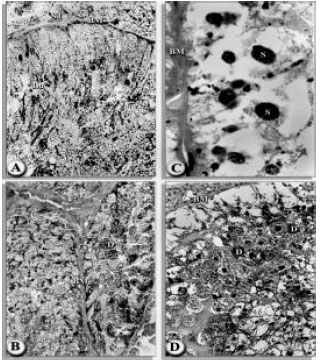
contained hematin granules and digestion appears to be progressing rapidly. From the 4th until the 6th day post-infection, the midgut revealed a severe damage in the basement membrane and digest cells. The food vacuoles have disappeared from the main cell present, the digest cell, which does not appear to be active in this stage (Fig. 5A-D). Secretory cells were present and basophilic cells, although present, were few.
Stienernema riobravae
A slight damage occurred in the basement membrane after 2 days of infection. A striking features was the presence of stem cells. Most of these cells seemed to be enlarging, some were dome-shaped which others became cuboidal as they expand. There are, in fact, all stages of cells intermediate between stem cells and digest cells. Some cells contained pale eosinophilic granules in the luminal bordered which appear to be small food vacuoles. The tick is feeding slowly and host blood cells and tissue fluid were seen in the lumen (Fig. 6A-D). Secretory cells were still enlarging and appear to be stretching towards the lumen.
Stienernema carpocapsae DD136
Many mitotic divisions are characteristic of this stage and digest cells apparently were abundant. Some of these cells were small and have empty cytoplasm, others were larger and have started ingesting the blood meal and contain small food vacuoles. Digest cells were either attached to the basal lamina or were free in the lumen and contained numerous hematin granules. A few intermediate stages exist with a combination of food vacuole and small hematin granules (Fig. 7A-D). Secretory cells were still enlarging and appeared to be stretching towards the lumen.
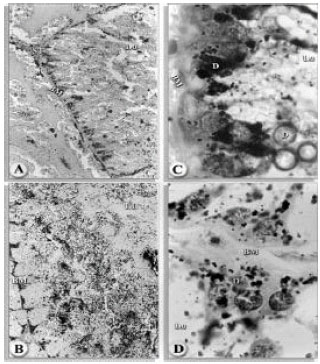 | |
| Fig. 3A-D: | Transverse sections of the midgut of H. dromedarii infected with Heterorhabditis indicus RM1 nematode; (A) After 2nd day of infection (160x). (B) After 4th day of infection (160x). (C) After 6th day of infection (1000x) and (D)After 6th day of infection (1000x) basement membrance (BM), digest cell (D), stem cell (ST), lumen (LU) Fig. |
 | |
| Fig. 4A-D: | Transverse sections of the midgut of H. dromedarii infected with H. bacteriophora HP88 nematode; (A) After 2nd day of infection (160x). (B) After 4nd day of infection (1000x). (C)After 6th day of infection (160x) and ( D)After 6th day of infection (1000x) basement membrance (BM), digest cell (D), hematin (H), lumen (LU) |
 | |
| Fig. 5A-D: | Transverse sections of the midgut of H. dromedarii infected with H. bacteriophora Eg1 nematode; (A) After 2nd day of infection (160x). (B) After 4th day of infection (160x). (C) After 6th day of infection (1000x) and (D)After 6th day of infection (1000x). Basement membrance (BM), digest cell (D), lumen (LU), stem cell (ST), secretory cell (S) |
 | |
| Fig. 6A-D: | Transverse sections of the midgut of H. dromedarii infected with Steinernema riobravae nematode; (A) After 2nd day of infection (160x). (B) After 4th day of infection (160x). (C) After 6th day of infection (1000x) and (D) After 6th day of infection (160x). Basement membrance (BM), digest cell (D), lumen (LU), stem cell (ST), secretory cell (S) |
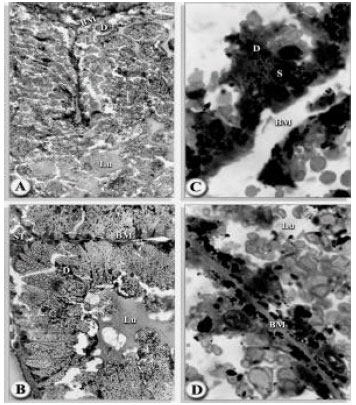 | |
| Fig. 7A-D: | Transverse sections of the midgut of H. dromedarii infected with S. carpcapsae DD 136 nematode; (A) After 2nd day of infection (160x) (B) After 4th day of infection (160x) (C) After 6th day of infection (1000x) and (D) After 6th day of infection (1000x).basement membrane (BM), digest cell (D), lumen (LU), stem cell (ST), secretory cell (S) |
Ultrastructure of Secretory Cells in H. dromedarii gut
Light microscopical studies have shown that, the digest cells, develop through a series of stages to mature cells packed with endosomes which are filled with host cells taken in from the gut lumen by pinocytosis and phagocytosis as suggested earlier (Agbede and Kemp, 1987). Furthermore, there are three generations of these digest cells during feeding of female ticks. Of the two secretory cell types present during the feeding, one (S2) appears to transform after feeding into basophilic cells which probably are quite different from its precursor. The female tick infected with H. bacteriophora HP88 used for electron microscopy had many maturing secretory cells of type 2 (S2), while S. riobravae infected ticks had many secretory cells of type 1 (S1) in the gut.
Secretory Cell Type 1
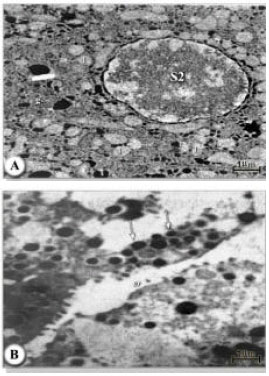
The S1 cells occurred singly, made close contact with the digest cells and were found in all regions of the gut. These cells (Fig. 8A) had a columnar to cuboidal shape, a slightly convex luminal surface and sparsely distributed microvilli which were much fewer than the S2 cells or the digest cells. The nucleus occupied the central part of the S1 cells and the cytoplasm was packed with homogenous densely-staining round granules. The cytoplasm also contained patches of cisternae of Rough Endoplasmic Reticulum (RER) which were concentrated around the nucleus. Mitochondria were scattered throughout the cytoplasm, but were absent towards the apical parts of the cells and there were no hematin granules or extensive infoldings of the basal lamina characteristic of other cell types. The possibility of another cell types that had not been observed by light microscopy is noted in S. riobravae infected ticks. These cells were seen in the gut caeca and were smaller than the S1 cells and contained numerous small dense granules in the cytoplasm. The luminal border of the cells had some thick cytoplasmic projections as well as micrvilli and the cell cytoplasm had patches of cisternas of RER (Fig. 8B).
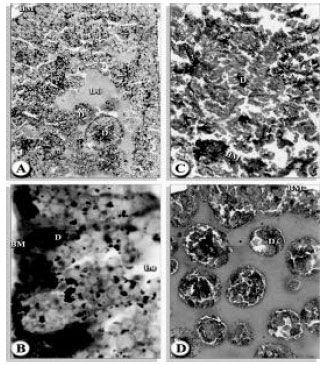
 | |
| Fig. 8A,B: | A) Electron micrograph of midgut of female H. dromedarii showing secretory cells type 1 (S1); secretory granules (g); digest cell (d); microvilli (mv); lumen (lu); mitochondria (m); distended rough endoplasmic reticulum (er). There were no pinocytic vesicles in these cells. B) Digest cells (d); pinocytic vesicles (pv); microvilli (mv); lumen (lu). The electron dense bodies in these cells lack the marginal vesicles shown in Fig. (A) |
 | |
| Fig. 9: | A, B Electron micrograph of midgut of female H. dromedarii showing secretory cell type 2 (S2); granules (g); rough endoplasmic reticulum (er); basal lamina, (bl); lipid-like granules, (l); arrows indicate glycogen deposits |
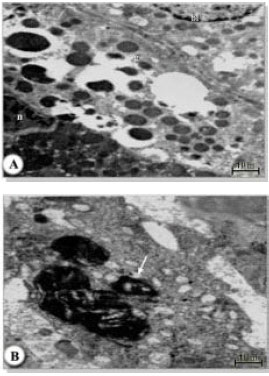 | |
| Fig. 10A,B: | Electron micrograph of midgut of female H. dromedarii showing secretory cells type 2 (S2); A) At the level of the glycogen and lipid accumulation. Nucleus (n); secretory granules (g). B) Serectory cell type 2 (S2); granule with broad projection (arrow) |
 | |
| Fig. 11A,B: | Electron micrograph of midgut of female H. dromedarii showing secretory cells type 2 (S2). A) Apical granules with electron lucent material. B) Basophilic cell developing from secretory cell type 2; remnant S2 secretory granules (rg); poorly organized rough endoplasmic reticulum (er);l hemolymph space (h); basal lamina (bl) |
Secretory Cell Type 2
Ultrastructurally, the luminal border of the cells had numerous closely spaced and uniformly sized microvilli, but no pinocytic vesicles. The cytoplasm was filled with well organized cisternae of RER whose vesicles were swollen (Fig. 9A and B). Large lipid-like granules and glycogen deposits formed accumulations above and below the nucleus but there were no hematin granules (Fig. 10A and B). Adjacent to the nucleus, there were small, parallel, flattened vesicles of the Golgi complex. The basal region of the cell had numerous infoldings of the plasma membrane forming a labyrinth, which extended deeply into the cell (Fig. 10A and B).
Development of Secretory Cell Type 2 to Basophilic Cells
The first stage of transition occurred during the period of rapid tick feeding as demonstrated in H. bacteriophora HP88 infected ticks. In the first stage, the basal portion of the cells had enlarged and the infolding of the basal labyrinth extended deeply into the cells, but the RER had not yet been recognized. In the next stage, after engorgement, further organization occurred, but the origin of the cells could still be traced because some of them had a few remaining S2 granules. The cells further enlarged and extended towards the lumen, then maintaining contact with the digest cells which were also expanding into the lumen as they ingested more of the blood meal. In the final stage of transformation, new, electron-dense, granules were synthesized. The granules occurred in aggregates and their ultrastructure and position in the cells was different from the granules which had formed earlier. By the fourth day of engorgement, many secretory granules had accumulated in the cells. Small infoldings of the basal membrane were found, but no typical structures associated with exocytosis of granules (Fig. 11A and B).
DISCUSSION
Studies on the midgut of ticks are important for the reason that it is the site for the development of blood born parasites transmitted by the ticks and secondly the midgut and its epithelial lining are a major physical barrier between the tick and the host defence mechanisms. The basic histology and ultrastructure investigations of the midgut epithelial cells during the digestion of the blood meal in ticks has been previously reviewed (Coons et al., 1982, 1986; Rosell-Davis and Coons, 1989). The present study of infected replete female H. dromedarii with five strains of entomopathgenic nematodes, (S. carpocapsae DD136, S. riobravae, H. bacteriophora HP88, H. indicus RM1 and H. bacteriophra Eg1), showed a highly significant damage in the midgut epithelial cells and the lesions increased by increasing the time of exposure. A significant difference was obviously noticed between Steinernematid and Heterorhabditid strains. The histopathological studies revealed a highly damaging effects with different species of Heterorhabditids (HP88, RM1 and Eg1). These results are in agreement with previous studies (Samish et al., 1999, 2000; Glazer et al., 2001).
Balashov (1972) described three basic cell types in the gut of H. asiaticum, namely, reserve cell, digest cell and secretory cell. He proposed that all gut cells have a common origin, the undifferentiated reserve cell being the initial type from which the other cells differentiate. In addition, secretory and digestive cells are irreversibly specialized. Nevertheless, one exception was noted and he observed a few secretory cells containing large hemoglobin food vacuoles, i.e., secretory cells transforming to digestive cells. Other investigators also believed that the three different cell types listed above comprise the midgut of ticks (Agbede and Kemp, 1985). Still others believe that the gut wall consists of one cell type, the form and structure of which may vary during different digestion phases and that these cells perform secretory and food digestion functions (Khalil, 1971).
In the present study, the presence of differentiated cells in all the midgut regions, as seen from serial sections, suggests that digestive processes appear to occur throughout the length and diameter of the midgut and its diverticulum as suggested earlier by Balashov (1972). Grandjean (1983) observed similar activity in the midgut of O. moubata. Similarly, Agyei and Runham (1995) found the midgut of B. microplus and R. appendiculatus to show synchronous cellular activity, which is likely to result from a trigger for differentiation to occur. The observation of a change in the activity of midgut digest cells on the arrival of the blood meal suggests that the presence of blood either mechanically or chemically may initiate the appearance of cells in the digest series. Mechanical stretching of the midgut with the arrival of the blood meal has been suggested (Szabo and Bechara, 1999) as a trigger for secretion. However, the absence of cells of the digest sites in some parts of the midgut epithelium suggests that parts of the midgut can function independently of other parts and that perhaps it is under some forms of local control.
The midgut digest cells has been considered to have a common origin (Tatchell, 1964; Guirgis, 1971; Balashov, 1972) and to be derived from the stem cells. We did not find any evidences to reject this. The morphological appearance of the cells in the present study are in agreement with the observations of Agbede and Kemp (1985) for B. microplus, Walker and Fletcher (1987) for R. appendiculatus and Agyei and Runham (1995) for both species.
The difficulty in defining the morphological nature of the midgut epithelial cells has contributed to the generation of multiple names for the same cell type. In the present work the names used with a few modification, are those of Agbede and Kemp (1985) and particularly of Walker and Fletcher (1987). In addition being suggested in the present work is the empty digest cells, in fact Agyei and Runham (1995) described some apparently empty-looking cells of the midgut of B. microplus and R. appendiculatus. The description for the empty digest cell in the present work is similar to the sessile digest cell of Walkar and Fletcher (1987). It is yet to be demonstrated whether the empty digest cells take further part in the digestion of the blood meal. However, the discharge of their hematin contents into the lumen may allow the cells to prepare itself for further intracellular digestion. Koh et al. (1991) have also observed similar activity in H. longicornis.
The accumulation of RNA in stem cells on differentiation, does not justify distinguishing one of the types of midgut digest cells as a basophilic cell, as it is not a distinct morphological entity. The transformation of the prodigest through sessile, residual sessile, detached to spent digest cells was accompanied by decreasing amounts of RNA in that order which, could reflect a reduction of their synthetic capacity. This loss of RNA with the transformation of the prodigest cells suggests that the various cells are in reality the different phases of a single cell type; the digest cell. This seems to reinforce the suggestion that the cells in the digest series are very likely to be a single cell type undergoing different phases of activity (Simo et al., 2004; Inayoshi et al., 2004).
In their ultrastructural study of the gut in H. asiaticum and O. papillipes, Balashov and Raikhel (1976) found one cell type that synthesized and secreted granules. However, the results from ultrastructural study on H. dromedarii showed that the granules of S1 and S2 secretory cells were different and the two cell types could sometimes be found together in one caecum. The cells differed from each other in size, shape and ultrastructurel. This S1 cell cytoplasmic organelles were virtually depleted by the time the granules matured, whereas the S2 cells still retained their organelles although they were in the process of being organized. This confirmed the evidence from light microscopy that there are two distinct secretory cell types in the gut of H. dromedarii.
All the secretory cells, including the basophilic cells, developed in close association with digest cells. Since digestion did not take place in the lumen and the excretory cells did not appear to ingest or digest material themselves, some of the metabolites may have entered by an unusual route, that is, directly from the digest cells rather than from the hemolymph. There was extensive membranous contact between the secretory cells and the digest cells and the hemolymph, but no specialization for transfer of materials across the membrane were found (Simser et al., 2004; Mosqueda et al., 2004).
There has been considerable speculation on the origin and transformation of gut cells in ticks. Tatchell (1964) was sceptical about the existence of distinct secretory cells in A. persicus. He suggested that after differentiation from the stem cells, synthesis and secretion of granules occurred and the cells then had the potential to transform to digest cells. In H. dromedarii, we did not find any evidence of secretory cells which transformed to digest cells, only secretory cells S2 which transformed into another secretory cell type (basophilic cells) after engorgement. On the other hand, Balashov (1972) studying O. papillipes, suggested that the secretory cells having completed their function, soon degenerated. In B. microplus, different studies suggested that S1 cells differentiated from the stem cells (Agbede and Kemp, 1985, 1987), completed their secretory function and disintegrated. The fate of the S2 cells however was not discussed and remained unsolved.
The origin of the secretory S2 cells also appeared to be from the stem cells (Agbede and Kemp, 1987) but the release of granular material and their subsequent transformation to basophilic cells was quite different from the S1 cells and was as complex as the transformation of the cells in H. dromedarii. The similarity in the ultrastructure of the S2 cells and the basophilic cells, particularly the presence of residual S2 granules in the basophilic cells, is further evidence for the origin of the basophilic cells.
After the ticks dropped from the host, the rough endoplasmic reticulum (RER) of the basophilic cells became organized and the cells synthesized new granules which were different from the S2 granules. The ultrastructure of these cells was very similar to the cells in ovipositing R. sanguineus which synthesized vitellogenin (Coons et al., 1982). The site of exogenous vitelloginin synthesis in the gut and fat body of ticks has been discussed by Diehl et al. (1982). It is likely that the material secreted by the gut basophilic cells in H. dromedarii is also vitellogenin. As suggested by Agbede and Kemp (1987) in B. microplus the gut material may be secreted into the extracellular spaces of the basal infolding which are in contact with the hemolymph. In the present study, it could be suggested that the membrane band granules possibly are released into the extracelluar spaces where their contents dissolve, or material could be released by exocytosis. The mechanism of secretion from the basophilic cells is as yet unknown.
REFERENCES
- Agbede, R.I.S. and D.H. Kemp, 1985. Digestion in the cattle-tick Boophilus microplus: Light microscope study of the gut cells in nymphs and females. Int. J. Parasitol., 15: 147-157.
CrossRefDirect Link - Agbede, R.I.S. and D.H. Kemp, 1987. Ultrastructure of secretory cells in the gut of the cattle-tick Boophilus microplus. Int. J. Parasitol., 17: 1089-1098.
CrossRefDirect Link - Agyei, A.D. and N.W. Runham, 1995. Studies on the morphological changes in the midguts of two ixodid tick species Boophilus microplus and Rhipicephalus appendiculatus during digestion of the blood meal. Int. J. Parasitol., 25: 55-62.
CrossRefDirect Link - Agyei, A.D., I.V. Herbert and N.W. Runham, 1991. Histochemical localization of acid phosphatase and non-specific esterase in the midgut of two tick species B. microplus and R. appendiculatus as determined by light microscopy. Parasitol. Res., 77: 629-634.
CrossRefDirect Link - Amosova, L.I., 2000. [An electron microscopic study of Borrelia in the body of the female ixodid tick Ixodes persulcatus]. Parazitologiia, 34: 234-240, (In Russian).
PubMed - Chinery, W.A., 1964. The mid-gut epithelium of the tick Haemaphysalis spinigera Neumann 1897. J. Med. Entomol., 1: 206-212.
CrossRefDirect Link - Coons, L.B., B.I. Tarnowski and D.D. Ourth, 1982. Rhipicephalus sanguinius: Localization of vitellogenin synthesis by immunological methods and electron microscopy. Exp. Parasitol., 54: 331-339.
CrossRefDirect Link - Glazer, I., E. Alekseev and M. Samish, 2001. Factors affecting the virulence of entomopathogenic nematodes to engorged female Boophilus annulatus ticks. J. Parasitol., 87: 808-812.
Direct Link - Grandjean, O., 1983. Blood digestion in Ornithodorus moubata Murray sensu stricto Walton females (Ixodoidea: Argasidae) II. Modifications of midgut cells related to the digestive cycle and to the triggering action of mating. Ann. Parasitol. Hum. Comp., 58: 493-514.
PubMed - Guirgis, S.S., 1971. The subgenus Persicargas (Ixodoidea, Argasidae, Argas) 13. Histological studies on A. (P.) arboreus Kaiser, Hoogstraal & Kohls. J. Med. Entomol., 8: 648-667.
CrossRefPubMedDirect Link - Hughes, T.E., 1954. Some histological changes which occur in the gut epithelium of Ixodes ricinus females during gorging and up to ovipostion. Ann. Trop. Med. Parasitol., 48: 397-404.
PubMedDirect Link - Inayoshi, M., H. Naictou, F. Kawamori, T. Masuzawa and N. Ohashi, 2004. Characterization of Ehrlichia species from Ixodes ovatus ticks at the foot of Mt. Fuji, Japan. Microbiol. Immunol., 48: 737-745.
Direct Link - Khalil, G.M., 1971. Biochemical and physiological studies of certain ticks (ixodoidea). Incorporation of tritiated tyrosine in the digestive system of nymphal Argas (Persicargas) arboreus (Argasidae). Ann. Entomol. Soc. Am., 64: 1149-1154.
CrossRefDirect Link - Kocan, K.M., E.F. Blouin, M.S. Oidherney, P.L. Claypoll, M. Samish and I. Glazer, 1998. Entomopathogenic nematodes as a potential biological control method for ticks. Ann. N. Y. Acad. Sci., 849: 355-364.
CrossRefDirect Link - Koh, K., T. Mori, S. Shiraishi and T.A. Uchida, 1991. Ultrastructural changes of the midgut epithelial cells in feeding and moulting nymphs of the tick Haemaphysalis longicornis. Int. J. Parasitol., 21: 23-36.
CrossRef - Mosqueda, J., A. falcon, J.A. Alvarez, J.A. Ramos, L.F. Oropeza-Hernandez and J.V. Figueroa, 2004. Babesia bigemina sexual stages are induced in vitro and are specifically recognized by antibodies in the midget of infected Boophilus microplus ticks. Int. J. Parasitol., 34: 1229-1236.
Direct Link - Rosell-Davis, R. and L.B. Coons, 1989. Relationship between feeding, mating, vitellogenin production and vitellogenesis in the tickDermacentor variabilis. Exp. Applied Acarol., 7: 95-105.
CrossRefDirect Link - Samish, M. and I. Glazer, 1991. Killing ticks with parasitic nematodes of insects. J. Invert. Parasitol., 58: 281-282.
PubMedDirect Link - Samish, M., E. Alekseev and I. Glazer, 1999. Efficacy of entomopathogenic nematode strains against engorged Boophilus annulatus females (Acari: Ixodidae) under simulated field conditions. J. Med. Entomol., 36: 727-732.
PubMedDirect Link - Samish, M., E. Alekseev and I. Glazer, 2000. Biocontrol of ticks by entomopathogenic nematodes. Ann. N. Y. Acad. Sci., 916: 589-594.
Direct Link - Simo, L., P. Kocakova, M. Slavikova, M. Kubes and V. Hajnicka et al., 2004. Dermacentor reticulatus (Acari: Ixodidae) female feeding in laboratory. Bologia Bratislava, 59: 655-660.
Direct Link - Simser, J.A., K.R. Macaluso, A. Mulenga and A.F. Azad, 2004. Immune-responsive lysozymes from hemocytes of the American dog tick Dermacentor variabilis and an embryonic cell line of the Rocky Mountain wood tick D. andersoni. Insect Biochem. Mol. Biol., 34: 1235-1246.
Direct Link - Szabo, M.P.J. and G.H. Bechara, 1999. Sequential histopathology at the Rhipicephalus sanguineus tick feeding site on dogs and guinea pigs. Exp. Applied Acarol., 23: 915-928.
PubMedDirect Link - Tatchell, R.J., 1964. Digestion in the tick, Argas persicus, (Oken). Parasitology, 54: 423-440.
CrossRefDirect Link - Walker, A.R. and J.D. Fletcher, 1987. Histology of digestion in nymphs of Rhipicephalus appendiculatus fed on rabbits and cattle naive and resistant to the ticks. Int. J. Parasitol., 17: 1393-1411.
CrossRefDirect Link