
J. P. Pandey
Department of Animal Sciences, School of Life Sciences, University of Hyderabad, Hyderabad-500046, India
R. K. Tiwari
Department of Zoology, R.N. Govt. PG. College, Gyanpur- 221304, S.R.N. Bhadohi, India
Dinesh Kumar
Department of Zoology, Banaras Hindu University, Varanasi-221, India
Journal of Entomology
Year: 2008 | Volume: 5 | Issue: 2 | Page No.: 113-121







ABSTRACT
Total and differential haemocyte counts following chilling, heating and ganglionectomy were studied in fifth instar larvae of Danais chrysippus. Chilling caused a decline in the number of haemocytes reaching a lowest level of about 57% in 48 h old larvae. Heating on the other hand elicited an increase of blood cells up to 30-50%. The ganglionectomy up to second abdominal ganglion caused a gradual reduction in total haemocyte count up to 93%. The relative percentage of different haemocyte types showed much variation. While prohaemocyte percentage increased both after heating and ganglionectomy but decreased after chilling; the percentage of plasmatocytes on the other hand declined in all experimental stages. The other cell types viz., granulocytes, spherulocytes, adipohaemocytes and oenocytoids also showed some variation in their counts. In addition, the aforesaid stresses adversely affected the structure of haemocytes causing their vacuolization, breaking of plasma membrane and fragmentation of nucleus and cell organelles leading to cell death. These stresses are thought to affect haemocytes through Median Neurosecretory Cells (MNSCs) mediated hormonal disturbance.
PDF Abstract XML References Citation
How to cite this article
J. P. Pandey, R. K. Tiwari and Dinesh Kumar, 2008. Temperature and Ganglionectomy Stresses Affect Haemocyte Counts in Plain Tiger Butterfly, Danais chrysippus L. (Lepidoptera: Nymphalidae). Journal of Entomology, 5: 113-121.
DOI: 10.3923/je.2008.113.121
URL: https://scialert.net/abstract/?doi=je.2008.113.121
DOI: 10.3923/je.2008.113.121
URL: https://scialert.net/abstract/?doi=je.2008.113.121
INTRODUCTION
Role of temperature in the life of insects is well known (Chapman, 1998; Sehnal et al., 2003; Solensky and Larkin, 2003). Temperature may have great effects on haemolymph, which contains several types of haemocytes with varied functions (Ribeiro and Brehelin, 2006). The haemolymph cells are known to play a major role in developmental processes during which they disassociate and rebuild metamorphosing tissues, though have been much studied in relation with different temperature regimes but the reports of various authors on their counts did not show any unanimity. While some of the workers (Tiwari and Shukla, 2000; Pandey, 2004) reported a decrease in Total Haemocyte Count (THC) at low temperature, few others (Rosenberger and Jones, 1960; Shapiro, 1979; Tiwari and Shukla, 2000) found an increase at higher temperature and still others found no change in cell number at lower temperature (Arnold, 1952; Rosenberger and Jones, 1960). Likewise, little and contradictory information are available on effect of temperature both on Differential Haemocyte Count (DHC) and the haemocyte morphology (Arnold, 1952; Behera et al., 1999; Pandey et al., 2003). Furthermore, the Ventral Nerve Cord (VNC) mediated regulation of haemocyte density is also less studied (Tiwari et al., 1997). The present study was, therefore, conducted to investigate the effects of chilling, heating and ganglionectomy (removal of ganglion) on THC, DHC as well as on the changes in haemocytes’ structure, if any, in the fifth instar larvae of D. chrysippus that feed voraciously on the leaves of Ak plant Calotropis gigantea known for its immense medicinal value. The insect under investigation poses a great threat to its host plant.
MATERIALS AND METHODS
Insects
The early larval instars of plain tiger butterfly, Danais chrysippus Linn. (Lepidoptera-Nymphalidae) were collected from Calotropis gigantea from the field in February and September. These larvae were reared on fresh leaves of the said plant in a BOD incubator at 27±1°C, 75±5% RH and 16L: 8D photoperiod in laboratory. Fifth instar larvae from the culture were subjected to various treatments as described below:
Chilling and Heating
Newly emerged (0 h old) to 108 h old fifth instar larvae were exposed to a chilling temperature of 4-6°C for 2 h by placing them directly on the ice cubes in a fridge. For heating, 24 and 96 h old larvae were put in an incubator at 50°C for an hour. Total Haemocyte Count (THC) and relative percentages of different types (prohaemocytes, plasmatocytes, granulocytes, spherulocytes, adipohaemocytes, oenocytoids and vermicytes) of haemocytes were calculated (vide infra).
Ganglionectomy
Fifth instar larvae (more than 24-h-old) of different body weight were narcotized by simply submerging them in water; these were subjected to surgical manoeuvering by the method described earlier (Shukla et al., 1993). In one group, second Thoracic Ganglion (TG2) and first and second Abdominal Ganglia (AG1 and AG2) were removed individually while in the second group, third and fourth Abdominal Ganglia (AG3 and AG4) were extirpated. In third group, only the sixth Abdominal Ganglion (AG6) was taken out. In controls, the ganglia were pulled but not removed (sham operated). The post-surgical treatments were also similar to the methods applied earlier (Shukla et al., 1993). THC were performed at designated periods (Table 3) till prepupa, pupa formation or mortality of the operated larvae whichever was earlier. The DHC, however, was performed at 72 h after ganglionectomy.
Haemocyte Counts
Total Haemocyte Count (THC)
Haemolymph (10 to 12 haemolymph determinations each) from oozing cut ends of prolegs of 4-5 larvae were drawn in a pretreated capillary with phenyl thiourea and diluted. The haemocytes were counted on a standard blood-cell counter under phase-contrast microscope. The mean number of circulating haemocytes per mm3 was calculated according to Jones (1962). Methods of permanent preparations of haemolymph smear and staining were adopted from Pandey and Tiwari (2004).
Differential Haemocyte Count (DHC)
It was made by counting at least 200 cells of different categories selected from random areas of stained smears of 10 insects from both the experimental and control groups separately and the percentage of different cell types was calculated. The data were subjected to the statistical analysis by using Student’s t-test.
RESULTS
Effect of Chilling and Heating
On THC and DHC-The total count of haemolymph cells declined to about 28% in the larvae subjected to chilling at the age of 24 h but this reduction reached the lowest level of about 57% in 48 h old larvae compared to controls (Table 1). Contrary to this, heating caused 1.3-1.5 fold increase in THC. The quantitative differences in haemocyte were observed more during heating than chilling temperature. DHC or haemocyte profile showed higher variation in the larvae exposed to these two temperature regimes (Table 2). While Prohaemocytes (PRs) and Plasmatocytes (PLs) percentages decreased in 24 h old chilled larvae, a little rise in the counts of Granulocytes (GRs), Spherulocytes (SPs), Adipohaemocytes (ADs ) and Oenocytoids (OEs) was recorded. But in 96 h old chilled larvae, the percentage of all cell types except PLs exhibited an increase (Table 2). In heat-exposed larvae, on the other hand, PRs, SPs and OEs rose in their number both in 24 and 96 h old larvae with a decline in other cell types.
On Cell Structure
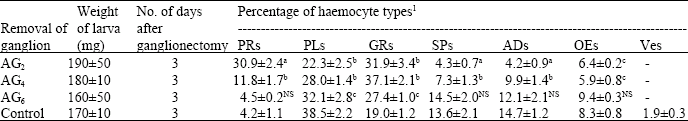
While chilling induced a little clumping of haemocytes (Fig. 1 and 2), heating caused damage of most of the cell types. Loss of cytoplasmic compactness (Fig. 3) loss of pseudopods in PLs, vacuolization, breaking of plasma membrane, fragmentation of nuclei and disintegration of sub-cellular organelles leading to cell death are some of the features observed in haemolymph smears of heat-exposed larvae.
Effect of Ganglionectomy
On THC and DHC
The effect of removal of various ganglia on THC is shown in Table 3. The removal of AG2 caused a gradual decrease in total cell number reaching the lowest level of 1020±190 (93%) and 2095±96 (86%) respectively depending on the larval body weight.
| Table 1: | Effect of chilling and heating on total haemocyte count in fifth instar larvae of Danais chrysippus* |
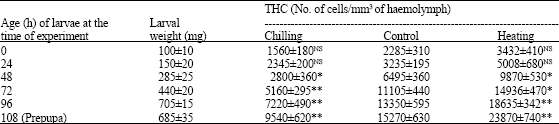 | |
*: Values represent mean±SD of 10-12 haemolymph determinations, each drawn from 4-5 larvae NS: Not significant; p-values: *<0.05; **<0.01 | |
| Table 2: | Effects of chilling and heating on differential haemocyte count in fifth instar larvae of Danais chrysippus |
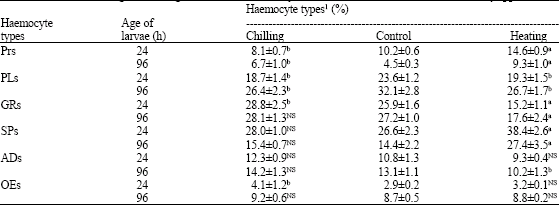 | |
1: Values represent mean±SD of 10-12 haemolymph determinations, each drawn from 4-5 larvae, NS: Not Significant; p-values: a<0.01; b<0.05 | |
| Table 3: | Effect of ganglionectomy on total haemocyte count in D. chrysippus |
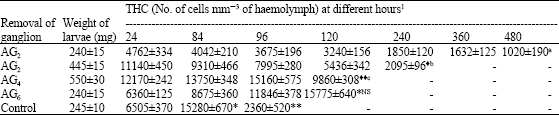 | |
Ë:Died as larva; * Prepupa; ♦Ë: LPI; **: Pupa, 1: Values represent mean±SD of 10-12 haemolymph determinations, each drawn from 4-5 larvae, NS: Not Significant; p-values: a, b<0.001; c<0.01 | |
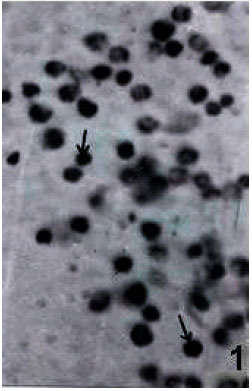 | |
| Fig. 1: | Blood smear showing small rounded PRs (arrows) and other haemocyte types x 675 |
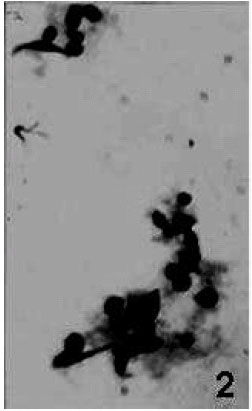 | |
| Fig. 2: | Clumping of haemocytes after exposure to chilling temperature x 450 |
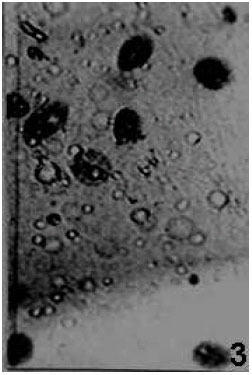 | |
| Fig. 3: | Loss of cytoplasmic compactness in haemocytes after exposure to heating temperature x 450 |
| Table 4: | Effect of ganglionectomy on differential haemocyte count in D. chrysippus |
 | |
1: Values represent mean±SD of 10-12 haemolymph determinations, each drawn from 4-5 larvae, NS: Not significant; p-values: a: <0.001; b<0.01; c<0.05 | |
These calculations were made taking the late fifth instar (prepupa) cell count as control. The larvae having a body weight of 240±15 mg survived for about 21 days whereas those with 445±15 mg survived for 10 days only. Larvae with a body weight of 460±25 mg and having their AG4 removed showed an increase in THC attaining a peak of 15160±575 in prepupa stage similar to that recorded in controls. Since ganglionectomised larvae of this group developed into Larval-Pupal Intermediates (LPIs) with a delay of about 2 days, the THC recorded was about 4 times more, compared to newly moulted control pupae. The removal of AG6 also led to an increase in THC despite a 1 or 2 day delay in moulting compared to control larvae. The newly ecdysed pupae of this experimental group have more or less the same haemolymph cell number as the control ones.
The haemocyte profile or DHC performed 3 day after ganglionectomy is shown in Table 4. The PR-percentage increased to about 7.5 times more and reached a maximum of 30.7% in experimental larvae devoid of their AG2. This rise in their number results from increased mitotic divisions of these cells. The number of PLs, on the other hand, revealed a decreasing trend. GRs also rose in test insects while SPs and ADs decreased with a very little fluctuation in OEs number. VEs were not traced in the test group since prepupa stage in which these cells appeared, were not formed by the time the haemolymph was pooled from these larvae.
On Cell Structure (Morphology)
Removal of ganglia viz., TG2, AG1 and AG2 affected the contour of different cell types. While PLs were seen to lose their cytoplasmic prolongations called pseudopods, the GRs appeared with 3 or 4 large granules on their periphery (Fig. 4).
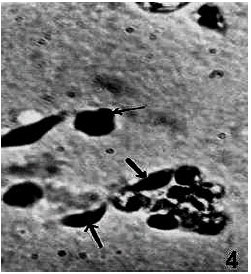 | |
| Fig. 4: | Loss of pseudopods in PLs (thick arrows) and appearance of large peripheral granules (thin arrow) in GR following removal of AG2. x 675 |
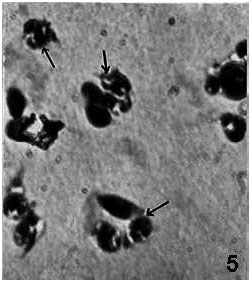 | |
| Fig. 5: | A number of cellular globules (arrows) comprising mostly PLs and GRs seen in blood smear after AG2 removal x 675 |
A large number of cellular globules perhaps phagocytic capsules constituted mostly by PLs and GRs were found abundantly in blood smear (Fig. 5). Such globules were rarely seen in the larvae having their ganglia removed between AG2 to AG6 but never in controls. Vacuolization of cells and breaking of the cellular organelles as well as nucleus took place leading to disintegration of cells.
DISCUSSION
Present results reveal that there are six types of haemocytes in D. chrysippus as largely met in Lepidoptera (Ribeiro and Brehelin, 2006). These haemocytes reveal adverse effects of stresses (chilling, heating and ganglionectomy- vide Tables), which might be taken akin to toxic stress as shown for Dysdercus koenigii (Tiwari et al., 2006). Such changes in haemocyte population in D. chrysippus larvae under altered physiological conditions are seen in other lepidopterans too (Gupta, 1979; Lackie, 1988; Sujatha and Gupta, 1991; Tiwari and Shukla, 2000; Pandey et al., 2003). Certain reasons could be assigned to these changes. For example, (I) loss of body fluid as a result of desiccation following increase in temperature, (ii) clumping of haemocytes or their detachment from tissue surfaces due to chilling or heating, respectively and (iii) change in the Mitotic Index (MI) of haemocytes, which in turn maintains the abundance of these cells in circulation in haemolymph. Presumably the effects of both external and internal stresses are known to be realized directly by the Median Neurosecretory Cells (MNSCs) of the brain, which are the site of synthesis and release of Prothoracicotropic Hormone (PTTH) leading to activation of the Prothoracic Glands (PTG). The variations in THC and DHC in experimental larvae under stress conditions might be caused due to the modified proprioception being conveyed to the brain, which thereby alters the PTTH release and hence the activity of PTG and its hormone. The importance of PTG and its hormone in production and differentiation of haemocytes is well documented (Hoffmann, 1970; Rao et al., 1984; Tiwari and Shukla, 2000). The modified proprioception following ventral nerve cord transection has also been reported to modify the MI of circulating haemocytes in Papilio demoleus (Tiwari et al., 1997).
The fall and surge in THC due to clumping or detachment of haemocytes respectively following chilling and heating and formation of cellular globules after ganglionectomy are presumed to be the various immune responses adopted by the experimental larvae to meet the challenges generated by these stresses. The role of haemocytes in immune reactions has recently been reviewed by Ribeiro and Brehelin (2006). Further, the haemocyte profile also varies a great deal in the stressed larvae of D. chrysippus. The multifold increase in PR-percentage despite overall reduction in THC especially in larvae devoid of their AG2 indicates the role of this cell type in maintaining high levels of haemocytes in circulation. This supports their stem cell nature as it has been reported Gupta (1985) Pandey and Tiwari (2004) and they could be responsible in providing strong tool to combat the stresses. Likewise the variations in counts of other cell types are dependent on various physiological functions ascribed to them as described by Pandey and Tiwari (2005) following VNC transection in D. chrysippus larvae.
The morphological changes in haemocytes leading to their disintegration observed under stress conditions match with the cellular changes as reported in Papilio demoleus (Pandey et al., 2003). These stresses might exert their adverse effects on MNSCs and could cause haemocytes disintegration similar to apoptosis occurring during hatching and moulting triggered by certain hormones being secreted from MNSCs (Chapman, 1998) eventually causing death.
The results do indicate that the haemocytes are temperature sensitive. Perhaps the PRs are most sensitive candidate inter alia haemocytes and seem, to be overall responsible for the normal physiological functions of the haemolymph. The ganglionectomy, on the other hand, points towards the fact that somehow hormone (ecdysteroids) titer is altered that affect the haemocytes number and hence function. It might be possible that in AG4 ganglionectomized larvae, the ecdysteroids titer is raised perhaps through raised THC causing larval-pupal moult. In addition, AG2 removal causes increase of PRs and GRs with an overall reduction in THC, which are presumably brought about by low ecdysteroids titer. One of the startling findings is that AG2 removal prolonged the larval span probably by interrupting their feeding thereby raising JH titer as reported for starved larvae of Galleria mellonela (Memmel et al., 1988). Thus the functions of AG2 and AG4 appear to be contradictory. On the other hand TG2, AG1 and AG2 might be responsible for the maintenance of normal morphology of the haemocytes via some neurohormones secretion. However, these could be ascertained only after further hormonal assay (RIA/ELISA etc). It appears that by manipulating the haemocytes one could control the development of D. chryssipus thereby checking their pest status in the field.
ACKNOWLWDGMENTS
RKT is thankful to Prof. A.N. Pandey, Principal and Department of Zoology, BHU, for providing laboratory facilities and to the University Grants Commission, New Delhi for financial assistance. Authors express their gratitude to anonymous referees for their critical evaluation of the manuscript.
REFERENCES
- Arnold, J.W., 1952. The haemocytes of the Mediterranean flour moth, Ephestia kuhniella Zell.(Lepidoptera: Pyralididae). Can. J. Zool., 30: 352-364.
CrossRefDirect Link - Chapman, R.F., 1998. The Insects: Structure and Function. 1st Edn., Cambridge University Press, Cambridge.
Direct Link - Hoffmann, J.A., 1970. [Endocrine regulation of production and differentiation of hemocytes in an orthopteran insect: Locusta migratoria migratoroides]. Gen. Comp. Endocrinol., 15: 198-219, (In French).
PubMedDirect Link - Jones, J.C., 1962. Current concepts concerning insect hemocytes. Am. Zool., 2: 209-246.
CrossRefDirect Link - Memmel, N.A., A. Ray and A.K. Kumaran, 1988. Role of hormones in starvation-induced delay in larval hemolymph protein gene expression in Galleria mellonella. Roux's Arch. Dev. Biol., 197: 496-502.
CrossRefDirect Link - Pandey, J.P. and R.K. Tiwari, 2005. Feeding, brain implantation and 20-hydroxyecdysone treatment reverses the effect of starvation and ventral nerve cord severance on haemocyte counts in larvae of the plain tiger butterfly Danaus chrysippus (Lepidoptera: Nymphalidae). Int. J. Trop. Insect Sci., 25: 295-300.
CrossRefDirect Link - Rao, C.G.P., A. Ray and P.S. Ramamurty, 1984. Effect of ligation and ecdysone on total haemocyte count in the tobacco caterpillar, Spodoptera litura (Noctuidae: Lepidoptera). Can. J. Zool., 62: 1461-1463.
CrossRefDirect Link - Ribeiro, C. and M. Brehelin, 2006. Insect haemocytes: What type of cell is that? J. Insect Physiol., 52: 417-429.
CrossRefDirect Link - Rosenberger, C.R. and J.C. Jones, 1960. Studies on the total blood cell counts of the southern armyworm larvae, Prodenia eridania (Lepidoptera). Ann. Entomol. Soc. Am., 53: 351-355.
CrossRefDirect Link - Solensky, M.J. and E. Larkin, 2003. Temperature induced variation in larval coloration in Danaus plexippus (Lepidoptera: Nymphalidae). Ann. Entomol. Soc. Am., 96: 211-216.
Direct Link - Shukla, R.S., R.K. Tiwari and D.P. Agrawal, 1993. Effect of ganglionectomy on the larval body from and metamorphosis of the Lemon-butterfly, Papilio demoleus L.(Lepidoptera). Zool. Jb. Physiol., 97: 277-283.
Direct Link - Sujatha, P.S. and A. Dutta-Gupta, 1991. Feeding and 20-hydroxyecdysone treatment reverses the effect of starvation on total haemocyte count in larval forms of Corcyra cephalonica (Lepidoptera). J. Stored Prod. Res., 27: 245-247.
CrossRefDirect Link - Tiwari, R.K., R.S. Shukla and A. Mishra, 1997. Effect of sectioning of ventral nerve cord on the total haemocyte count in lemon-butterfly, Papilio demoleus L. Indian J. Exp. Biol., 35: 1378-1380.
Direct Link