
Amit Sethi
Department of Entomology and Nematology, University of Florida, P.O. Box 110620, Gainesville, FL 32611-0620, USA
M.S. Bons
Department of Entomology and Nematology, University of Florida, P.O. Box 110620, Gainesville, FL 32611-0620, USA
V.K. Dilawari
Department of Entomology, Punjab Agricultural University, Ludhiana-141 004, India







ABSTRACT
Whitefly, Bemisia tabaci (Gennadius) is a serious pest of many economic crops through out the world. Farmers depend highly on insecticide usage to manage this pest. In this study, we investigated the capacity of field collected whitefly populations from different crops to develop resistance against imidacloprid, bifenthrin and fenvalerate by selecting up to 8 generations. Selection pressure was given by exposing the adults to insecticides using the treated leaf discs in the Petri dishes at the dosages sufficient to give 60-80% mortality. After 8th generation, strains selected with these three insecticides exhibited 21.90, 7.12 and 4.13-fold increase in tolerance for the three insecticides, respectively. The realized heritability of insecticide resistance was very high in imidacloprid-selected strains compared to bifenthrin and fenvalerate-selected strains. This indicated a high level of risk in the field populations for the development of resistance to imidacloprid compared to bifenthrin and fenvalerate. The mode of inheritance of insecticide resistance was found to be controlled by nearly completely recessive more than one gene.
PDF Abstract XML References Citation
How to cite this article
Amit Sethi, M.S. Bons and V.K. Dilawari, 2008. Realized Heritability and Genetic Analysis of Insecticide Resistance in Whitefly, Bemisia tabaci (Genn.). Journal of Entomology, 5: 1-9.
DOI: 10.3923/je.2008.1.9
URL: https://scialert.net/abstract/?doi=je.2008.1.9
DOI: 10.3923/je.2008.1.9
URL: https://scialert.net/abstract/?doi=je.2008.1.9
INTRODUCTION
The sweet potato whitefly, Bemisia tabaci (Gennadius) is an important pest of cotton, vegetables and ornamentals worldwide (Servín-Villegas et al., 2001). It was known as secondary pest since 1920’s (Azab et al., 1971; Byrne et al., 1990). But in the last two decades, it has increased its host and geographical range and attained the status of a primary pest (Dittrich et al., 1985; Schuster, 2003). Plant damage is caused by both direct feeding on the plant and transmission of plant diseases (Byrne et al., 1992; Sivasupramaniam and Watson, 2000).
Resurgence of whitefly population has been reported from many countries (Dittrich et al., 1985, 1990; Prabhaker et al., 1985; Horowitz, 1986; Jayaraj et al., 1986). The climatic factors (Jayaraj et al., 1986; Byrne et al., 1992), cropping practices (Byrne et al., 1992) and the use of insecticides (Ditrich et al., 1990; Wool and Greeenberg, 1990; Mochizuki, 1994; Ortega, 1998) were responsible for the resurgence. Application of sub-lethal doses of insecticides has been reported to increase reproduction in many insect species (Ball and Su, 1979; Chelliah and Heinrich, 1980) including B. tabaci (Dittrich et al., 1985; Byrne et al., 1992). An over-reliance on broad spectrum insecticides plays an important role in the outbreak of this insect due to the development of insecticide resistance (Dittrich et al., 1990; Wool and Greeenberg, 1990; Mochizuki, 1994; Ortega, 1998). The proportion of resistant individuals in a population increases due to the evolutionary adaptation of insects to their chemical environment which eliminates genetically susceptible individuals (Roush and McKenzie, 1987). Resistance in several insects to commonly used insecticides is widespread (Georghiou, 1990) and continued use of conventional insecticides is likely to intensify selection for resistance in the field thereby aggravating the problem of resistance. For resistance management tactics to be effective, resistance must be detected in its early stages (Roush and Miller, 1986) and early detection necessitates testing of large number of individuals at each location where resistance is suspected.
The purpose of our study was to investigate status of resistance in B. tabaci to insecticides, such as imidacloprid, bifenthrin and fenvalerate and to elucidate the genetics of resistance in the natural populations from Punjab state of India. Information about the genetic basis of resistance can facilitate efforts to detect and monitor resistance, to assess the risk of resistance, to model the evolution of resistance and to delay resistance development in B. tabaci.
MATERIALS AND METHODS
Insecticides
Commercial formulations of test insecticides Confidor 200 SL (imidacloprid), Bifenthrin 10 WP (bifenthrin) and Fenval 20 EC (fenvalerate) were used. Serial dilutions of the insecticides were prepared in distilled water.
Plants and Insects
Potted plants of cotton, Gossypium hirsutum var. F846 were raised (free of insecticides) in large insect proof cages inside screen-house. Five to six weeks old cotton plants were used for rearing whiteflies. Wooden cages (45x45x60 cm) lined with insect-proof mesh on two sides and sliding glass pans to facilitate operations on the other two sides were used to raise whiteflies in a screen-house. The top of cage was also lined with fixed glass pan for ambient lighting and temperature conditions. A large number of genetically diverse populations of whitefly (> 10, 000 adults) were collected in 2001 from different crops in Punjab, India and a mixed colony was established. Subsets of the colony were used to establish 4 strains in separate screen-houses selected with (1) imidacloprid, (2) bifenthrin, (3) fenvalerate and (4) the unselected founder strain which was not exposed to any insecticide. Each strain was reared in 2-3 similar cages. Care was taken to minimize the risk of contamination by hanging yellow sticky traps.
Bioassays and Selection Procedure
The bioassay method was adapted from Dittrich et al. (1985) with slight modifications to determine the tolerance levels of adults of each strain to imidacloprid, bifenthrin and fenvalerate. Cotton leaf discs were dipped in aqueous solution of formulated materials for 10 seconds and allowed to dry for 1 h. To maintain the turgidity after drying, the treated leaf disc was placed in a plastic Petri dish lid with 4.0 cm diameter containing a thin layer of agar. Twenty to thirty (unsexed) whitefly adults were transferred in the Petri dish containing leaf disc. The Petri dish was kept upside down with lid inverted over the other half of the dish so that the insects attach themselves to the underside of the leaf disc in their regular feeding position. Ventilation was provided by mesh-covered openings drilled on the sides of the Petri dish. These Petri dishes were held at 27.0±2.0°C with a photoperiod of 12:12 (L:D) and mortality was recorded after 24 h. Control tests were conducted with cotton leaf discs without any insecticide treatment. A series of 5 doses was used for each insecticide with 5 replications. Concentration-mortality regression was worked out using probit analysis package POLO-PC (Le-Ora Software, 1987 based on Finney, 1971). The selection pressure equivalent to 60-80% mortality was applied through 9 generations.
Data Analyses
Slope and LC50 values based on concentration-mortality regression were compared between generations to monitor the development of resistance. Two LC50 values were considered significantly different if their 95% fiducial limits did not overlap. Tolerance ratio was computed as a ratio between LC50 of unselected strain population and LC50 of the insecticide-selected strain. Resistance risk assessment was made by calculating realized heritability (h2) values as described by Tabashnik (1992):
Where:
| R | = | Response to selection |
| S | = | Selection differential (Hartl, 1988; Falconer, 1989) |
Response to selection (R), the difference in mean phenotype between offspring of the selected parents and the parent generation before selection (Falconer, 1989) was estimated as:
Where:
| Final LC50 | = | LC50 of the offspring after n generations of selection |
| Initial LC50 | = | LC50 of the parental generation before n generations of selection |
The difference between LC50s was calculated on a logarithmic scale because the logarithm of tolerance was assumed to be normally distributed while the numerator of above equation for R estimates the cumulative responses to selection over n generations.
The selection differential (S), the difference in mean phenotype between the selected parents and the entire parental generation (Hartl, 1988) was estimated as:
Where:
| i | = | Intensity of selection |
| σp | = | Phenotypic standard deviation |
Intensity of selection (i) was estimated from p, which is the percentage of the population with values above the selection threshold (i.e. the percentage surviving selection) using Appendix of Falconer (1989), which is based on the properties of normal distribution.
The phenotypic standard deviation (σp) was estimated as the reciprocal of the mean of the estimated slopes of probit regression lines (Finney, 1971) from the parental selection before insecticidal selection (initial slope) and the offspring after ‘n’ generations of selection (final slope).
The number of generations (G) required for a 10-fold increase in LC50 was also calculated as the reciprocal of R.
The degree of dominance (of resistant component) was estimated separately for the three insecticide-selected strains with the following formula (Stone, 1968).
Where:
| D | = | Dominance of the examined character (resistance) |
| Y1 | = | Log10 of LC50 of F0 generation of the unselected check strain |
| Y2 | = | Log10 of LC50 of F8 generation of the insecticide-selected strain |
| Y3 | = | Log10 of LC50 of F1 generation of the insecticide-selected strain |
This formula will result in a value of -1 if the resistance is completely recessive, a value of 0 if there is no dominance and a value of +1 if the resistance is completely dominant.
The number of independent genes with additive effects that contribute to the expression of a trait (insecticide resistance) was estimated from the mortality data obtained in successive generations selected with three insecticides as per Raymond et al. (1987).
Another independent method given by Lande (1981) to estimate the number of genes with additive effects contributing to the expression of a quantitative trait (insecticide resistance) was:
Where:
| σ2 | = | Genetic variance of the insecticide-selected strain, estimated as (slope-1)2 |
| N | = | No. of generations |
RESULTS
Response of Whitefly to Imidacloprid, Bifenthrin and Fenvalerate
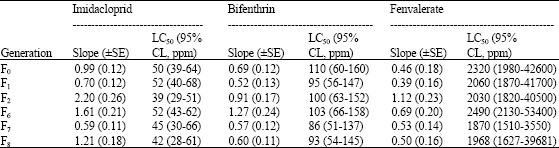
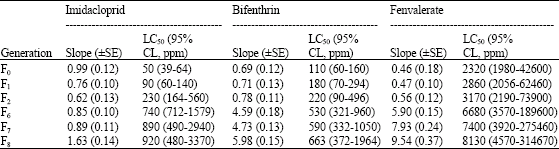
The LC50s of the unselected strain for imidacloprid did not show any significant change during the period of selection (Table 1) where as selection with imidacloprid at LC60-80 concentration for 8 generations produced an increase in LC50 (Table 2). Whiteflies did not show any appreciable change in the tolerance to imidacloprid during the first two generations (LC50 ranged from 50 to 90 ppm). Tolerance ratio during this period was 1.73 fold (Table 3). Minimal increase in LC50s was seen between generations F7 (890 ppm) and F8 (920 ppm, 1.03-fold). Beginning in the F2 generation, an upward trend continued, culminating in 21.90-fold tolerance ratio in the F8 generation with LC50 of 920 ppm. The results did not show appreciable increase in the values of slope in the successive generations in spite of continued selection pressure. The slope values related to imidacloprid ranged from 0.62 to 1.63.
LC50 values of the unselected strain to bifenthrin and fenvalerate also showed no perceptible change during the course of 8 generations. The LC50s values for bifenthrin and fenvalerate of unselected population were ranged invariably between 93-110 and 1870-2490 ppm, respectively in the successive generations (Table 1). Whitefly populations through 8 generations of selection achieved only moderate levels of increased tolerance (Table 2, 3). The LC50 of bifenthrin and fenvalerate-selected strains continued to increase over the generations and peaking in F8 (LC50 = 663 and 8130 ppm, TR = 7.12 and 4.13-fold, respectively). With the development of resistance over the generations, these strains showed a correspondingly increasing slope values when selected with bifenthrin (0.69-5.98) and fenvalerate (0.46-9.54).
| Table 1: | Susceptibility of unselected population of whitefly to different insecticides |
 | |
| Table 2: | Susceptibility of the insecticide-selected strain of whitefly to different insecticides |
 | |
| Table 3: | Tolerance ratio of whitefly strains selected with different insecticides in successive generations |
 | |
Resistance Risk Assessment
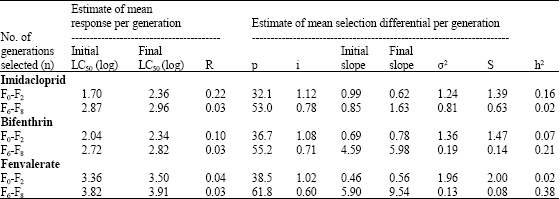
Estimated h2 to imidacloprid was 0.16 and 0.02 in the F0-F2 and F6-F8, respectively (Table 4). On the other hand, estimated h2 was 0.07 and 0.21 for the F0-F2 and F6-F8, respectively in case of bifenthrin and 0.02 and 0.38 for the F0-F2 and F6-F8, respectively in case of fenvalerate.
Projected Rates of Resistance Development
The number of generations (G) required for 10-fold increase in LC50 was estimated to be 39, 64 and 150 in case of imidacloprid, bifenthrin and fenvalerate-selected strains, respectively.
Degree of Dominance of Resistance Trait
The degree of dominance of the resistant trait in imidacloprid, bifenthrin and fenvalerate-selected strains was -0.60, -0.43 and -0.52, respectively. This indicates that insecticidal resistance against all the three insecticides is nearly completely recessive.
No. of Genes Involved in Resistance Trait
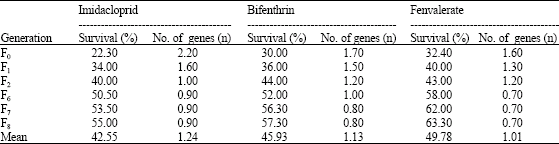
The number of genes involved in insecticide resistance against imidacloprid, bifenthrin and fenvalerate as per Raymond et al. (1987) was found to be more than 1 up to F2 generation and ultimately, resistance was controlled by single gene (Table 5).
With the use of Lande’s (1981) formula, the number of genes segregating across eight generations was estimated to be 4.85, 3.09 and 2.95 for the insecticide resistance against imidacloprid, bifenthrin and fenvalerate, respectively.
| Table 4: | Realized heritability (h2) of insecticide resistance in whitefly |
 | |
| Table 5: | Estimation of number of genes contributing to insecticide resistance in whitefly against different insecticides |
 | |
DISCUSSION
The low slope values of imidacloprid-selected strain indicate considerable heterogeneity in its response and also suggest a greater potential for the development of higher levels of imidacloprid resistance. On the other hand, the high slope values of bifenthrin-and fenvalerate-selected strains point to the establishment of highly homozygous populations with regard to bifenthrin and fenvalerate resistance. The increase in tolerance ratio in all the three insecticide-selected strains of whitefly over the generations indicates that insecticide resistance is at least in part genetically determined (Bloch and Wool, 1994) and is also additive in nature (Falconer, 1989).
The value of realized heritability obtained at the end of 3 generations (F0-F2) of imidacloprid-selected strain is very high (8 times) than those for F6-F8 generations and could indicate a high level of risk in the field populations for development of resistance to imidacloprid. The results also suggest that brief selection period (3 generations) may be sufficient to detect the potential for the development of resistance. Where as in case of bifenthrin-and fenvalerate-selected strains, the values estimated at the end of 3 generations (F0-F2) are 3 and 19 times less than those for F6-F8 generations, respectively. This reflects that high levels of resistance to bifenthrin and fenvalerate can only be realized after long periods of selections (8 generations) in the field populations of whitefly. One of the important uses of heritability estimates is the prediction of future response (Hartl, 1988; Falconer, 1989). The purpose of insecticide resistance studies is to predict the rate of development of resistance in response to insecticide application (Via, 1986; Firko and Hayes, 1991). Heritability estimate after one generation of selection is often a reliable approximation of the heritability of the trait in the parental population because laboratory environment has minimal effects (Tabashnik, 1991). As the mean response for the first three generations is high in case of imidacloprid, therefore risk for development of resistance is higher compared to than that in bifenthrin and fenvalerate. Almost 39 generations are required to develop 10-fold increase in LC50 of imidacloprid where as more than 64 and 150 generations are required in case of bifenthrin and fenvalerate, respectively.
Estimates of dominance in this study indicated that insecticide resistance is nearly completely recessive. This pattern could result from the additive inheritance of multiple genes (Keena and Granett, 1990). The inheritance of resistance according to Raymond et al. (1987) formula is controlled by multiple genes in the early generations under continuous selection pressure while single gene in the later generations. Calculations based on Lande’s (1981) formula suggest that insecticide resistance against imidacloprid, bifenthrin and fenvalerate is controlled by more than 1 gene. This can further be substantiated by the significant increase in tolerance ratios in response to the selection over the generations. Apparently, combinations of alleles responsible for tolerance which are not present in the parental generations, are produced in the succeeding generations (under continuous selection pressure) as a result of inbreeding among tolerant individuals as suggested by Bloch and Wool (1994). Roush and McKenzie (1987) noted that most significant cases of resistance are caused by allelic variants at one or two loci. They reasoned that multiple gene resistance is favored by laboratory regimes that select at moderate doses from small samples where as field applications that select at high doses from large populations favor single gene resistance by rare alleles. Without suitable genetic markers, use of bioassays to discriminate between inheritance mediated by a single gene with modifiers and a more complex mode of inheritance with multiple genes is generally difficult and is almost impossible with overlapping concentration-response lines (Tsukamoto, 1963).
In conclusion, although very high levels of resistance is attained by these laboratory-selected populations, yet these differences may not necessarily interpret to the reduction or loss of field performance of these insecticides (Denholm et al., 1984). We must apply caution in directly extrapolating results from laboratory experiments to field conditions. Field populations are usually more heterogeneous and exhibit more complex and diverse response to insecticide pressures. Interactions of environment, population structure and selection intensity greatly affect field response (Bloch and Wool, 1994). The evolution of insecticide resistance under field conditions could be delayed due to immigration of susceptible populations from other crops and the alteration of insecticides. The degree of insecticide resistance has been shown to reduce down when the insecticide pressure is removed (Servín-Villegas et al., 2001). However, many of the insecticides used to control whiteflies are rendered ineffective more easily because of the occurrence of cross-and multiple-resistance.
REFERENCES
- Ball, H.J. and P.P. Su, 1979. Effect of sublethal dosages of Carbofuran and carbaryl on fecundity and longevity of the female western corn rootworm. J. Econ. Entomol., 73: 873-876.
CrossRefDirect Link - Bloch, G. and D. Wool, 1994. Methidathion resistance in the sweetpotato whitefly (Aleyrodidae: Homoptera) in Israel: Selection, heritability and correlated changes of esterase activity. J. Econ. Entomol., 87: 1147-1156.
CrossRefDirect Link - Byrne, F.J., I. Denholm, A.L. Devonshire and M.W. Rowland, 1992. Analysis of Insecticidal Resistance in the Whitefly, Bemisia tabaci. In: Resistance 91: Achievements and Development in Combating Pesticide Resistance, Denholm, I., A.L. Devonshire and D.W. Hollomon (Eds.). Elsevier, London, pp: 165-178.
- Chelliah, S. and E.A. Heinrich, 1980. Factors affecting insecticide-induced resurgence of the brown Planthopper, Nilaparvata lugens1 on rice. Environ. Entomol., 9: 773-777.
CrossRefDirect Link - Dittrich, V., S.O. Hassan and G.H. Ernst, 1985. Sudanese cotton and the whitefly: A case study of the emergence of a new primary pest. Crop Protect., 4: 161-176.
CrossRefDirect Link - Dittrich, V., G.H. Ernst, O. Ruesch and S. Uk, 1990. Resistance mechanisms in sweetpotato whitefly (Homoptera: Aleyrodidae) populations from Sudan, Turkey, Guatemala and Nicaragua. J. Econ. Entomol., 83: 1665-1670.
CrossRefDirect Link - Falconer, D.S., 1989. Introduction to Quantitative Genetics. 3rd Edn., Longman Scientific and Technical, New York, USA Pages: 438.
Direct Link - Finney, D.J., 1971. Probit Analysis. 3rd Edn., Cambridge University Press, London, UK., pp: 76-80.
CrossRefDirect Link - Firko, M.J. and J.L. Hayes, 1991. Quantitative genetic analysis of larval resistance to cypermethrin in tobacco budworm (Lepidoptera: Noctuidae). J. Econ. Entomol., 84: 34-40.
CrossRefDirect Link - Horowitz, A.R., 1986. Population dynamics of Bemisia tabaci (Gennadius): With special emphasis on cotton fields. Agric. Ecosyst. Environ., 17: 37-47.
CrossRefDirect Link - Jayaraj, S., A.V. Rangarajan, S. Murugesan, G. Santhramj, S.V. Jayaraghovan and D. Thangaraj, 1986. Studies on the Outbreak of Whitefly, Bemisia tabaci (Gennadius) on Cotton in Rice in Tamil Nadu. In: Resurgence of Sucking Pests, Jayaraj, S. (Ed.). Proc. Natl. Symp. Tamil Nadu Agric., Coimbatore, India, pp: 103-115.
- Keena, M.A. and J. Granett, 1990. Genetic analysis of propargite resistance in Pacific spider mites and twospotted spider mites (Acari: Tetranychidae). J. Econ. Entomol., 83: 655-661.
CrossRefDirect Link - Lande, R., 1981. The minimum number of genes contributing to quantitative variation between and within populations. Genetics, 99: 541-553.
Direct Link - Prabhaker, N., D.L. Coudriet and D.E. Meyerdirk, 1985. Insecticide resistance in the sweetpotato whitefly, Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol., 78: 748-752.
CrossRefDirect Link - Raymond, M., N. Pasteur and G.P. Georghiou, 1987. Inheritance of chlorpyrifos resistance in Culex pipiens L. (Diptera: Culicidae) and estimation of the number of genes involved. Heredity, 58: 351-356.
CrossRefDirect Link - Roush, R.T. and G.L. Miller, 1986. Considerations for design of insecticide resistance monitoring programs. J. Econ. Entomol., 79: 293-298.
CrossRefDirect Link - Roush, R.T. and J.A. McKenzie, 1987. Ecological genetics of insecticide and acaricide resistance. Ann. Rev. Entomol., 32: 361-380.
CrossRefDirect Link - Schuster, D.J., 2003. Preference of Bemisia argentifolii (Homoptera: Aleyrodidae) for selected vegetable hosts relative to tomato. J. Agric. Urban Entomol., 20: 59-76.
Direct Link - Servin-Villegas, R., E. Troyo-Dieguez and J.L. Martinez-Carrillo, 2001. Wild hosts of Bemisia argentifolii bellows and perring in semiarid Northwest Mexico. Southwestern Entomol., 26: 239-244.
Direct Link - Stone, B.F., 1968. A formula for determining degree of dominance in cases of monofactorial inheritance of resistance to chemicals. Bull. World Health Organiz., 38: 325-326.
Direct Link - Tabashnik, B.E., 1991. Determining the mode of inheritance of pesticide resistance with backcross experiments. J. Econ. Entomol., 84: 703-712.
CrossRefDirect Link - Tabashnik, B.E., 1992. Resistance risk assessment: Realized heritability of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae), tobacco budworm (Lepidoptera: Noctuidae) and colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol., 85: 1551-1559.
CrossRefDirect Link - Tsukamoto, M., 1963. The log dosage-probit mortality curve in genetic researches of insect resistance to insecticides. Botyu-Kagaku, 28: 91-98.
Direct Link - Wool, D. and S. Greenberg, 1990. Esterase activity in whiteflies (Bemisia tabaci) in Israel in relation to insecticide resistance. Entomol. Exp. Applied, 57: 251-258.
CrossRefDirect Link