
Riffat Sultana
Department of Zoology, University of Sindh, Jamshoro, Pakistan
M. Saeed Wagan
Department of Zoology, University of Sindh, Jamshoro, Pakistan
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 5 | Page No.: 379-386







ABSTRACT
Hieroglyphus nigrorepletus I.Bolivar is reported as injurious pest of rice, sugarcane, wheat, maize and minor pest of millets and fodder crops in Pakistan. Presently a fair number of specimens both nymphs and adults were collected from various province of Pakistan. Present study was carried out in field as well as in laboratory. The hoppers go through six nymphal stages before becoming adult. The developmental rate was recoded 4.37±0.0, 5.02±0.14, 5.73±0.05, 4.79±1.20, 5.64±0.38 and 4.92±0.82 days for first to six instars, respectively. Hoppers usually emerge during July as soon as monsoon begins and adults appear in August-September in field. The total population of hopper was significantly higher during the hottest months of the year i.e., July, August, September and October. The sex ratio is in the favour of females. Similarly the green colour form was more prominent in females as (64.53%) than males (35.46%). Morphological description of various nymphal instars and measurement of different body parts has been discussed. In addition to this, present finding also suggests that mortality percentage of hoppers is increased with an increased in their population density in laboratory.
PDF Abstract XML References Citation
How to cite this article
Riffat Sultana and M. Saeed Wagan, 2007. Life History and Economic Importance of Hieroglyphus nigrorepletusBolivar (Hemiacridinae: Acrididae: Orthoptera) from Pakistan. Journal of Entomology, 4: 379-386.
DOI: 10.3923/je.2007.379.386
URL: https://scialert.net/abstract/?doi=je.2007.379.386
DOI: 10.3923/je.2007.379.386
URL: https://scialert.net/abstract/?doi=je.2007.379.386
INTRODUCTION
The acridid grasshopper Hieroglyphus nigroreletus I.Bolivar is reported as a major pest of rice, sugarcane, wheat, maize and as a minor pest of millets and fodder crops in Pakistan and India (Roonwall, 1978). It was in 1950 that this pest was reported by Bhatia to be causing extensive damage to millets in Ajmer Marwara state and in the adjoining areas of Rajastan about 75% of the village covering an area of 112.707 acres of maize and jowar in Ajmer-Marwara state was reported to be infested with this grasshopper. Ghouri and Ahmed (1960) reported a medium-size swarm of H. nigrorepletus approximately 500 specimens from Karachi, Malier and smaller swarm was also examined from Bela, Thatta and adjoining areas of Hyderabad. Collection of such greater number of macropterous form would be warning on potential future plague. Qadri (1971) recorded irregularly or regularly-occurring outbreaks in Pakistan caused by 4 orthopteran species including H. nigrorepletus. Moizuddin (2001) reported some incidence of H. nigrorepletus in desert area of Lasbela Balochistan. Recently Wagan and Riffat (2004 and 2005) and Riffat et al. (2007) carried extensive study on the distribution of H. nigrorepletus from Pakistan.
Despite numerous information on systemic status of this pest, an inadequate description of breeding habit of the H. nigrorepletus were given by Chaturvdi (1946) Srivastava (1956) Roonwall (1945, 1976a, 1976b and 1978) Pradhan and Peswani (1961) Rizvi and Khan (1970) Uvarov (1966 and 1977), Mason (1973) and Jotwani and Butani (1978) Siddiqui (1986 and 1989) from India, but their information is mostly concerned with copulation and oviposition, life cycle of this rice grasshopper pest is not fully understood. Its habitat, seasonal fluctuation and egg diapause vary from place to place depending on climatic and ecological conditions. It is therefore, felt necessary to study the life history of H. nigrorepletus from this region. Hoppers appear to be more epidemic than adults. Taking into consideration the economic status of the immature stages of H. nigrorepletus, present study has been carried out under laboratory condition as well as in field.
MATERIALS AND METHODS
Collection of Samples
The nymphs of H. nigrorepletus were randomly collected from the fields having rice, maize, jowar, wheat, millets and fodder crops with the help of traditional insect hand-net (8.89 cm in diameter and 50.8 cm in length) and by hand picking during the year 2003 to 2005 in the months of July-November.
Rearing and Maintenance of Insect
Nymphs were raised up to the adulthood under laboratory condition (25°-23/N 68°-24/ E), where the Relative humidity 26 to 61% and temperature ranged between 28±2 and 39±2°C. Housing and feeding conditions were identical for all the nymphs in cages as well as in separate jars. Fresh leaves of Zea mays were cut from field, immersed in water and served as food to experimental insects.
To determine the number and duration of nymphal instars in the laboratory, the first intar hoppers were reared singly in one-pound glass-jars. The jars were given separate number. The hoppers were reared on Z. mays. The body parts of the hoppers were measured with Vernier-Calliper except the antennae and first instar. The antennae and first instar hoppers were measure by Ocular Square Graph and the number of segments of antennal and sex- ratio of instars was carried out under Stereoscopic Dissecting Binocular Microscope (SM90). All the measurements were taken in millimeter (mm).
To study the effect of population density on the survival of hoppers; three cages of same size (Length 8½, Width 5 ½ cm) were used. In each cage 25, 50 and 100 hoppers were kept. They were fed on Z. mays. Each day, the observations were taken and the dead hoppers were removed from the cages. Beside this, the number of moulted hoppers was recorded. Longevity of insects was also recorded.
Statistical Analyses
Data obtained from experimental insects were subjected to one-way analysis of variance (ANOVA) with repeated measures and significant means were determined using Duncan’s New Multiple Range test (DNMRT). The computer programme (SPSS 10.0 soft ware) was used to compare the means of the various treatments.
RESULTS
Numbers of Molt
Present study suggests that H. nigrorepletus go through six nymphal stages before becoming adult and there is only one generation per year. The average duration was recorded 4.37±0.0, 5.02±0.14, 5.73±0.05, 4.79±1.20, 5.64±0.38 and 4.92±0.82 days for the first to six instars, respectively (Table 1) the resulting figures show that stage I took shorter time (4.37±0.0 days) whereas the time taken by stages II, IV and VI was almost equal. On the contrary, the duration of stage III and V was recoded prolonged than the rest of instars. However, the average duration was recorded (5.07±0.47 days) for all the nymphs. Beside, this survival period of hopper was recorded maximum (30.48 days) and minimum (23.30 days). However, majority of them could not complete the life cycle. Hardly 10% survived. There was no significant difference in the development rate of male and female hoppers. As mentioned above six nymphal instars were observed.
| Table 1: | The effects of Z. mays on nymphal developmental period of H. nigrorepletus |
 | |
| Mean in the same column followed by the same letter (s) are not significantly different from one another at 5% level of probability (DNMRT) | |
Morphological Description of Hopper Stages
First Instar
Eyes reddish brown, small brownish dots on the fastigium of vertex. Front and genae with certain reddish spots. Antennae 10-14 segmented (10 mostly in male), filiform flattened in the basal part and gradually narrowing and thin apically. Pronotum cylindrical; without marking. The lateral margin of prozona and metazoana slightly extended. Prozona is larger than metazoana. Meso and meta thorax without elytron and wing rudiment. Abdomen yellowish/green in color, covered with thin hairs. Hind femur yellowish green with brown stripes.Tibia yellowish green; with thin hairs and spines. Genitalia not well differentiated (Fig. 1a).
Second Instar
Eyes dark brown. Antennae with 14-20 segments (14 mostly in male). Frontal ridge with strong depression extended 0.7 mm in width and 4.5 mm length. Pronotum without marking. Elytron and wing rudiments small but distinct, pointing downward dorsally. Male genitalia chitinised, with a weak, obtuse notch at the posterior margine. In female the anterior and posterior ovipositor valves small but well marked; middle valves not developed. Abdomen yellowish/ green without marking. Tibia yellowish green cover with hairs and spines (Fig. 1b).
Third Instar
Antennae with 13-23 segments (13 rarely). Clypeus divided by a thin yellowish line. Blackish dots on the frontal ridge and gena. Frontal ridge with deep depression; pronotum without marking. Elytron and wing rudiments a little more developed and pointing downward. Male genitalia rounded and extending a little backward. In female the anterior and posterior ovipositor valves a little further developed but do not yet overlap the posterior valves; middle valves not yet clearly differentiated. Tarsi with blackish tip (Fig. 1c).
Fourth Instar
Antennae with 19-25 segments. Fastigium of vertex become large extended 1.4 mm in length and 2.8 mm in width. Brown dots on the meso-and-metanotum. Elytron and wing rudiments small and pointing downward. Male genitalia now pointed and extending over the following sternum. In female the anterior ovipositor valves overlapping the posterior ones; middle valves differentiated but concealed beneath the anterior valves. Cerci well developed and larger than supra annal plates (Fig. 1d).
Fifth Instar
Antennae with 21-25 segments (mostly 25, 17 rarely). Frontal ridge with slight depression Eyes pigment more prominent. Certain brown dots on the compound eyes. Prozona is slightly rounded than metazona. No brown dots on meso-and-metanotum. Elytron and wing rudiments a little more developed; have undergone an un-turning and now point upward; wing rudiments covering part of the first abdomen segment. Male genitalia further developed, extending beyond the hind margin of the following sternum. In female the ovipositor valves a little more developed (Fig. 1e).
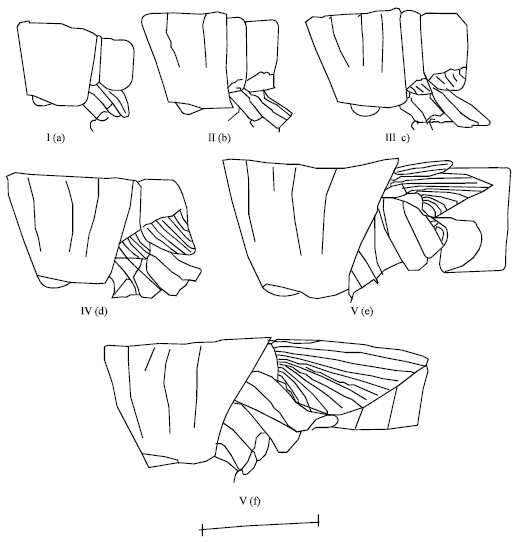 | |
| Fig. 1: | Growth of wing rudiments in different stages of H. nigrorepletus(Scale line = 2 mm) |
Sixth Instar
Antennae with 24-26 segments. The lateral side of pronotum with prominent yellowish color from where sulci developed. Four thin sulci beginning to appear at this stage laterally on the pronotum. Elytron and wing rudiments further developed and remain pointing upward; the former extending a little beyond the metathorax, the latter a little beyond the first abdominal segment. Male genitalia extending well beyond the hind end of the following sternum. In female the ovipositor valves extending backward a little more. Abdomen with dark brown and yellowish color (Fig. 1f).
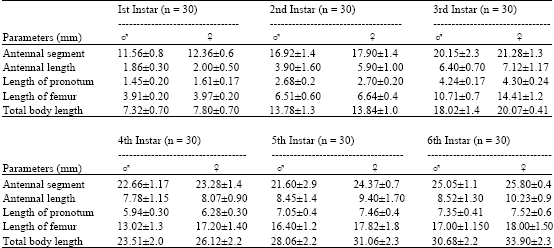
Measurement to Distinguish Various Instar of H. nigrorepletus
Table 2 suggests that in first instar there was no significant difference in the total body length and length of femur in female than that of male while other all parameters were insignificant. In second instar antennal segments, length of pronotum and femur was significantly different in female compare with male. However in third instar there was no significant difference in the pronotum of male (4.24±0.17 mm) and female (4.30±0.24 mm). As far as fifth and sixth instars are concerned antennal segments, length of femur and total body length of female was significantly greater, excluding the length of pronotum that was equal in male and female.
| Table 2: | Measurements (mm) used to distinguish various instars of H. nigrorepletus |
 | |
| Table 3: | Sex ratios in green and brown colour forms in hopper (Stage II-VI) and adult, H. nigrorepletus as collected from field |
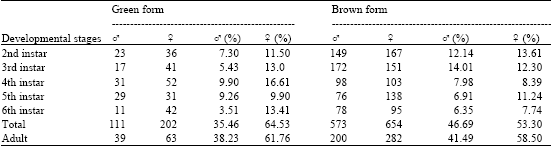 | |
| Note: A total of 2087 nymphs and 584 adults were examined to count the green and brown coloration | |
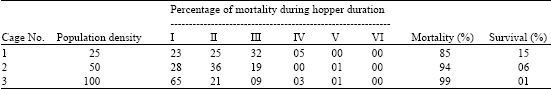
| Table 4: | Effect of population density of H. nigrorepletus hoppers on their survival |
 | |
Field Population Density
During the field survey it was noted that hoppers usually emerged from the mid July immediately after the first monsoon showers begin and adults appear in August-September the maximum activities of the pest were observed from mid-August to October in the field when the copulation and oviposition take place. The duration of the hoppers stage is recorded five to six weeks in field. However, their population beings to decline from first week of November with decrease in temperature.
Sex Ratio and Color Form among the Hoppers and Adult
Table 3 revealed that percentage of females was significantly higher (59.07%) as compared to males (40.92%). Moreover females of H. nigrorepletus was found extremely green than males.
Population Density in Laboratory
Mortality ratio was significantly higher in the earlier stages i.e., I-III (Table 4). However, mortality also occurred in the IV and V stages when the density of hopper was 50 and 100. The hoppers survived through III stage moulted to adult. It was investigated that survival percentage in case of above mentioned densities was (15, 06 and 01%), respectively in same size cages, the hoppers with different population densities were kept and were provided similar condition except variation in per individual hopper space. Present finding suggests that population density had greater effect on the survival percentage of the hoppers.
DISCUSSION
Existing literature shows that there is variation in the number of moult. Pruthi (1949) stated that H. nigrorepletus pass through 5 nymphal instars to become adult. Srivastava (1956) reported that H. nigrorepletus moult seventh time. Pradhan and Peswani (1961) observed that two out of eight nymphs reared at 26±1°C moulted 7 times instead of 6 as in all other cases. Roonwall (1976b) studied that there are 6 moult (excluding the intermediate moult) and 6 hopper’s stages excluding the vermiform larva. Occasionally, there is an extra moult in stage IV. Presently no extra instar was found in stage IV. However, present study suggests that this variation might be due to effect of different climatic conditions of the region. Presently the period between hatching and first moult and between previous and succeeding moulting was considered as the nypmhal duration.
Roonwal (1976b) gave inadequate description of hopper stages his illustration does not clarified the exact position of wings rudiments. Presently position of wing rudiments is well documented which is easy to follow distinguish between different instars. Small and inconspicuous wing rudiments are found in first Instar while Narayanan and Samuel (1954) stated that newly emerged hopper of H. nigrorepletus is practically wingless. One of the reasons for the irregularity of growth rate throughout the instars might be due to the different rates of growth between the various body parts and between instars in the same part. Previously actual mode of growth has been studied for only a few parts and organs of grasshopper (Katiyar, 1960; Richards and Waloff, 1954; Remaudhiere, 1954). There are, of course some individual variation within a species as well as different due to sex. As far as colourmorphism is concerned, explanation for this variation due to different ages and sexes of insect, under different culturing conditions, habitats and season of the region.
According to Roonwall (1945) H. nigrorepletus undergoes one generation each year. Eggs are laid in the soil during August and September and hatching takes place during the following June or July soon after the first monsoon showers. The present finding is slightly differ with Indian author for the reason that the monsoon rains occur in early summer in June and July in India in late summer in July and August in Pakistan, especially in the area of NWFP (Roonwall, 1945), also noted that the duration of the hoppers stage is three weeks, but in the present study it has been noted the duration of the hopper stages is five to six weeks, which is contrary to above findings. However, present study revealed that the rains in July are therefore important, because, if this month was dry, a large percentage of the eggs would fail to hatch. On the whole, early and uniformly distributed summer rains create favorable conditions for this species. Maximum population of hoppers as well as adults was reported in hottest months of the year it suggests that cold temperature discouraged the buildup of grasshoppers while the hot temperature encouraged it. The present study strongly recommends that control techniques are being applied to immure stage of insect because these are more epidemic than adults. However, the results of present study might be useful to forecast the exact hatching date and should be of benefit in avoiding or preventing any possible future outbreak.
ACKNOWLEDGMENTS
We wish to express our sincere gratitude to Prof. Dr. N.T. Narejo Department of Fresh water Biology and Fisheries, University of Sindh, Jamshoro for his kind guidance for statistical analysis and valuable comments on manuscript. This study was financial supported by Pakistan Science Foundation Islamabad for research grants (Project No. PSF S-SU/Bio 338).