
T. Srinivasan
Department of Agricultural Entomology, Tamil Nadu Agricultural University,
Coimbatore, 641 003, Tamil Nadu, India
C. Durairaj
Department of Pulses, Tamil Nadu Agricultural University,
Coimbatore, 641 003, Tamil Nadu, India
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 5 | Page No.: 371-378







ABSTRACT
Nineteen accessions of the wild plants including rice bean viz., Vigna umbellata Thunb. (Ohwi and Ohashi), a wild relative of the genus Vigna and other species belonging to Vigna were screened for their relative resistance to the bruchid, Callosobruchus maculatus F., a serious pest of stored pulses. The results showed that, accessions of V. umbellata were found to arrest the growth and development of C. maculatus, at grub stages in varying levels. Morphological characters like seed coat thickness and seed hardness were not found to be responsible for offering resistance to C. maculatus. The death of the grubs at the cotyledons was mainly due to the anti-nutritional factors in the cotyledons. Among the various biochemicals analysed, the trypsin inhibitors in the resistant accessions were in the order of 3-5 times higher (1576.12 to 3120.08 TIU g-1) than in the susceptible check (682.09 TIU g-1). Similarly the cysteine protease inhibitors were 7 times higher in the resistant accessions (2061.78 to 4923.62 CPIU g-1) compared to the susceptible CO 6 (686.77 CPIU g-1). The protein profile also showed the presence of low molecular weight proteins in the range of 14-29 kDa in the resistant accessions. The correlation between different anti-nutritional factors with that of index of suitability also showed a negative relationship. The role of these antinutritionals in relation to the C. maculatus infestation is discussed.
PDF Abstract XML References Citation
How to cite this article
T. Srinivasan and C. Durairaj, 2007. Biochemical Basis of Resistance in Rice Bean, Vigna umbellata Thunb. (Ohwi and Ohashi) Against Callosobruchus maculatus F. Journal of Entomology, 4: 371-378.
DOI: 10.3923/je.2007.371.378
URL: https://scialert.net/abstract/?doi=je.2007.371.378
DOI: 10.3923/je.2007.371.378
URL: https://scialert.net/abstract/?doi=je.2007.371.378
INTRODUCTION
Pulses constitute a major source of protein for the people living in developing countries like India, where the per capita consumption of animal protein is relatively low. These pulses are referred to as poor man’s meat and rich man’s vegetable (Singh and Singh, 1992). Wide array of pests including Helicoverpa armigera Hubner, Maruca vitrata Geyer, Clavigralla gibbosa Spinola, Melanagromyza obtusa Malloch, Mylabris sp. etc. attack the crop during vegetative and reproductive stages (Durairaj, 1999). However, the bruchid, Callosobruchus maculatus Fab. which infest the seeds at storage assume special significance as it is a very serious pest causing up to 100% storage loss. Infestation of these insects starts in the field and continues in storage where sometimes it causes total destruction of the seeds within a period of 3-4 months (Dongre et al., 1996; Hall et al., 1997; Sarikarin et al., 1999). The estimated losses due to bruchids in various pulses ranged from 30-40% within a period of six months and the post harvest seed losses due to bruchids can reach even 100% during severe periods of infestation (Mahendran and Mohan, 2002).
The present management options of physical methods do not provide the expected levels of control. Insecticides though effective, cause other problems including residual effects and are carried through the food chain leading to disastrous consequences. Thus, enormous focus is being given to host plant resistance in recent years. The wild plants belonging to the genus Vigna are a potent source of resistance as evidenced by several authors. Marconi et al. (1997) observed several species of Vigna including, V. vexillata, V. reticulata, V. luteola, etc. as resistant to bruchids. The antinutritional factors including trypsin inhibitors, tannins and phytic acid were found to be higher in these species. Ignacimuthu et al. (2000) have also stressed the importance of protease inhibitors including trypsin and chymotrypsin inhibitors in conferring resistance against C. maculatus.
However, more thrust is yet to be given for the utilization of wild relatives of mungbean for the management of bruchids. Earlier report showed that, an accession TC 1966 of Vigna radiata var. sublobata was found to be completely resistant to Callosobruchus chinensis (Fujii and Miyazaki, 1987). Identification of resistant donors from non-edible leguminous tree seeds like and Vigna bourneae (Gamble) was attempted by Ignacimuthu et al. (2000). In view of the various reports available for the presence of resistant sources in the wild plants, an attempt has been made to assess the level of resistance in Vigna umbellata to C. maculatus and the possible biochemical factors responsible for the same.
MATERIALS AND METHODS
Rearing for Insects
The bruchid species, C. maculatus used for this work has been obtained from a culture maintained continuously at the Biocontrol Unit of Tamil Nadu Agricultural University, Coimbatore following the procedure of Strong et al. (1968). Permanent culture of beetles was established on V. radiata (cv. CO 6) and once in a month subculturing was done in order to maintain a continuous culture. The insects were maintained at a temperature of 30±5°C and 70±5% r.h. throughout the period of study.
Seeds
Seventeen accessions of V. umbellata, a wild Vigna species, Vigna glabrescens and a cross, VRM Gg1 x V. sublobata were obtained from TNAU, Coimbatore and raised for a season to obtain sufficient seed lot for the experiment. A cultivar, CO 6 was used as check for the entire study.
Bruchid Development on Different Accessions
To measure the performance of C. maculatus on different accessions of V. umbellata, three replicates of 25 seeds of each accession was used. Three pairs of freshly emerged adults of C. maculatus were released into a polypropylene envelope containing the test seeds. The insects were allowed for oviposition for 72 h under no choice conditions (Gibson and Raina, 1972). After that period, the insects were removed and the number of eggs laid on each accession was recorded. The seeds were left undisturbed for a period of about 60 days and the number of adults emerging during this period was recorded on a daily basis for calculating the per cent survival to adult emergence (S), mean developmental period (T) and index of suitability (I).
Biophysical Parameters
The biophysical parameters viz., hundred seed weight, seed coat thickness and seed hardness of seeds of different accessions were recorded (Table 1) for assessing the presence of antixenotic mechanisms, if any.
Biochemical Analysis
Trypsin and chymotrypsin inhibitory activity was measured following the procedure of Kakade et al. (1969) and the cysteine protease inhibitory activity was measured following the procedure of Barrett (1981).
| Table 1: | Biophysical characteristics of wild Vigna species |
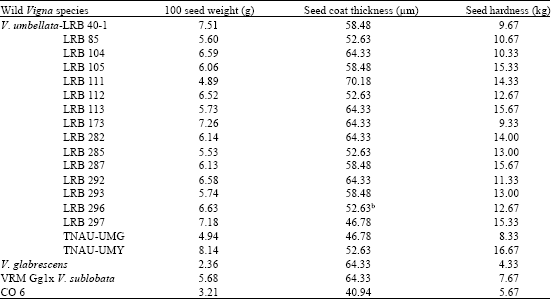 | |
They were expressed in terms of inhibitory units/g of seed materials. One unit of activity corresponds to that amount of trypsin/cysteine protease/chymotrypsin inhibitor which gives 50% inhibition of enzyme activity under experimental conditions. The inhibitory activity was expressed as trypsin/chymotrypsin/cysteine protease inhibitor units (TIU/CIU/CPIU) per g of sample.
Seed Protein Profile
To analyse proteins of promising Vigna species, SDS-polyacrylamide gel electrophoresis (SDS-PAGE) was performed according to the method of Laemmli et al. (1970). Proteins were extracted from the seeds with 0.1 M Tris-HCl buffer, pH 8.0 in a prechilled pestle and mortar for 15 min at 0-4°C. The extracts were centrifuged at 15000 g for 10 min at 4°C. The protein concentration in the extracts was adjusted in each well using five fold sample buffer and water in such a way that the same amount of protein was present per unit volume. Standard medium range molecular markers (Genei, Bangalore, India) were simultaneously run for comparison.
Statistical Analysis
All data were analyzed using the AGRES package and the level of significance was tested at p = 0.05. Correlation of different parameters was done using SPSS package.
RESULTS
The data on oviposition, adult emergence and developmental period showed significant differences. TNAU-UMY recorded the least number of eggs (11.67) and LRB 113 the maximum number (63.33) while the control CO 6 registered 33.33 eggs (Table 2). With regard to adult emergence on the 19 accessions tested, 15 accessions showed no adult emergence, the exceptions being LRB 113, LRB 296 and V. glabrescens. The percentage survival was also considerably lesser in these accessions when compared to CO 6 (28.41%). Further, those accessions which exhibited lesser per cent survival encountered a prolonged developmental period. The delay in developmental period was much pronounced in LRB 296 (38.67 days) as against CO 6 (30.90 days). Ultimately the index of suitability was lowest in LRB 113 (0.006) and was followed by LRB 282 (0.0097), LRB 296 (0.0181) and LRB 297 (0.0261) and highest in the check variety CO 6 (0.0469).
| Table 2: | Biological performance of Callosobruchus maculatus on different species of wild Vigna species |
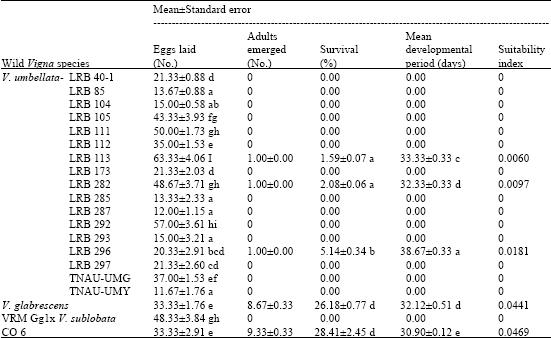 | |
| In a column, means followed by a common letter(s) are not significantly different by LSD (p = 0.05) | |
| Table 3: | Quantity of various antinutritional factors in the accessions of Vigna umbellata |
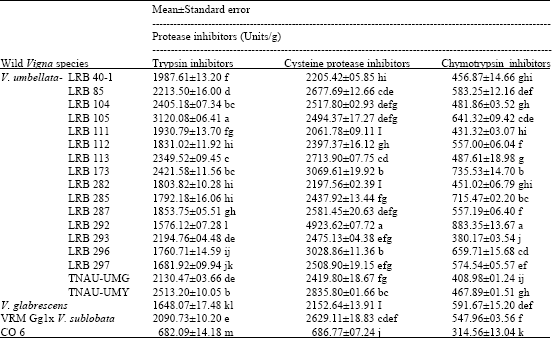 | |
| In a column, means followed by a common letter(s) are not significantly different by LSD (p = 0.05), Values represent mean of three replications; NS = Non significant | |
Following these bioassay studies biochemical analyses were made to further study the reason underlying low rates of survival in the wild accessions when compared to CO 6. Analyses of biochemical constituents in the wild Vigna accessions revealed that the protease inhibitors viz., trypsin, cysteine protease and chymotrypsin inhibitors were relatively higher in the wild Vigna species (Table 3). Among the accessions, significantly higher levels of trypsin inhibitor was noticed in LRB 105 (3120.08 TIU g-1) while a least quantity of 1648.07 TIU g-1 was noticed in V. glabrescens.
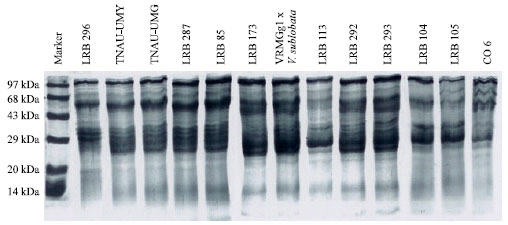 | |
| Fig. 1: | SDS-PAGE (Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis) profile of promising accessions of wild Vigna species) |
However the quantity of trypsin inhibitors was considerably lower in CO 6 (682.09 TIU g-1). Similar was the case with respect to cysteine protease inhibitors. The accession LRB 292 registered the maximum quantity of cysteine protease inhibitors (4923.62 CPIU g-1) which corresponds to a seven times higher quantity than CO 6 (686.77 CPIU g-1). Even LRB 111 which registered the least quantity of cysteine protease inhibitor among the accessions (2061.78 CPIU g-1) had three times higher quantity than CO 6. With respect to chymotrypsin inhibitors also, LRB 292 showed a maximum quantity of 883.35 CIU g-1 which was 2.8 times higher than CO 6 (314.56 CIU g-1).
The SDS-PAGE protein profile of the wild Vigna accessions revealed a moderate banding pattern in the lower molecular weight region (Fig. 1). A polypeptide band corresponding to an apparent weight of 14 kDa could be noticed in all the wild Vigna species except the check CO 6. The region between 14 and 29 kDa exhibited similar banding patterns in all the wild accessions while CO 6 was devoid of such a pattern. Further an intense banding corresponding to a molecular weight of 29 kDa was observed in all the wild Vigna species while CO 6 exhibited a less intense banding in these regions.
DISCUSSION
All the wild Vigna species used in the present study were preferred for oviposition by C. maculatus. However except four accessions (LRB 113, LRB 282, LRB 296 and V. glabrescens) all the wild accessions failed to support the emergence of adults. Even these four accessions were found to delay the developmental period of C. maculatus. The developmental period of C. maculatus in LRB 113, LRB 282, LRB 296 and V. glabrescens was 33.33, 32.33, 38.67 and 32.12 days, respectively. On the other hand C. maculatus grubs completed their life cycle in a relatively shorter span of 30.90 days in CO 6.
The biophysical factors like 100 seed weight and seed hardness had a highly significant and negative relationship with that of index of suitability (r = -0.62 and -0.47, respectively) (Table 4). However, these factors might not have contributed to the resistance in these seeds. Because, the seeds of V. umbellata were heavier than the green gram seeds and were having sufficient resources for a growing grub. Moreover, if seed hardness had a role, the insects should have had difficulty in penetrating the cotyledons. Thus, the assumption by Southgate (1979) that size and hardness of the seeds influencing the adult emergence does not hold good. The seed coat thickness also cannot be considered as factors conferring resistance as the grubs penetrated and reached the cotyledons in all the cases. Thus, the death of the grubs in the cotyledons might be due to the presence of toxic substances present in the seeds.
| Table 4: | Correlation of biophysical and biochemical parameters to index of suitability |
 | |
| *Significant at p = 0.05, **Significant at p = 0.01, NSNon significant | |
Earlier, Kashiwaba et al. (2003) opined that several accessions of V. umbellata completely inhibited the emergence of C. maculatus which support the present investigations. The cross, VRM Gg1 x V. sublobata showed complete resistance to C. maculatus though there was egg laying. An accession, TC 1966 of V. radiata var. sublobata was also found to be immune to C. maculatus (Fujii and Miyazaki, 1987).
Thus, the presence of antinutritional factors in the cotyledons of these resistant accessions is confirmed by the fact that all the wild Vigna seeds permitted penetration by the grubs, but failed to support a normal adult emergence Kashiwaba et al. (2003) witnessed mortality of first and second instar larva inside cotyledons of V. umbellata. Earlier Lale and Makoshi (2000) also observed larval mortality of C. maculatus in the seed coats of certain cowpea cultivars, when first instar grubs were trying to gain entry. Thus from the above discussions it is obvious that the only possible reason that could be attributed to the non-emergence of adults is the presence of toxins in the cotyledons. The findings of Kashiwaba et al. (2003) also reflected the above inference when they used artificial beans made of V. umbellata flour and found decreased adult emergence and prolonged mean developmental period of C. maculatus. Furthermore, the biochemical constituents having a negative correlation with that of index of suitability in the present studies supported the possible role of antinutritional factors for the non emergence of C. maculatus adults.
Significantly higher quantities of protease inhibitors was noticed in all the tested accessions when compared to CO 6. The role of protease inhibitors had been well documented by Shulke and Murdock (1983) who reasoned out that, protease inhibitors were capable of inhibiting the activity of digestive enzymes and reduced the quantity of proteins that can be digested. This leads to insufficient utilization of proteins by the insect and as a result the insect become weak with stunted growth and ultimately die.
Among the protease inhibitors, the trypsin inhibitor content in the wild Vigna species was considerably higher (1576.12 to 3120.08 TIU g-1) than in the susceptible check, CO 6 (682.09 TIU g-1). Moreover the trypsin inhibitor content had a highly significant and negative relationship at 1% level (r = -0.66**) stressing the importance of trypsin inhibitor as a potent antinutritional factor (Table 4). Saikia et al. (1999) reported a range of 2456 to 2534 TIU g-1 in different Vigna umbellata cultivars which backs the present findings. However, the role of trypsin inhibitor in bruchid resistance has been contradicted by Janzen et al. (1977) who observed that the diets incorporated with trypsin inhibitors were found to have little or no effect on C. maculatus. The accessions LRB 292 had higher amount of cysteine protease inhibitor and chymotrypsin inhibitor with low levels of trypsin inhibitor. The cysteine protease inhibitor also had a significant and negative relationship at 5% level with that of index of suitability (r = -0.48*). Hence, the resistance in this accession might have been governed by the trypsin and cysteine protease inhibitors alone. The role of cysteine protease inhibitor in conferring resistance to C. maculatus had been documented by many authors. Hines et al. (1991) observed that the soybean cysteine protease inhibitors inhibited the proteolytic activity of the gut extracts of C. maculatus. Koiwa et al. (1998) also observed that when soybean cysteine protease inhibitors were incorporated into a diet of C. maculatus, there was a prolonged developmental period and increased mortality in cowpea. Indepth investigations by Ryan (1989) revealed that, the bruchids have cysteine protease as the predominant digestive enzymes and hence the inhibition of these enzymes would be of greater importance in conferring resistance against C. maculatus. Moreover the cysteine protease inhibitors had a significant and negative correlation with that of index of suitability, which affirms that cysteine protease inhibitors had an active role as an antinutritional factor. The amount of chymotrypsin inhibitors in wild Vigna species was found to be generally lower than that of trypsin inhibitors. A similar phenomenon was observed in non edible legumes by Ignacimuthu et al. (2000).
SDS-PAGE profile done for promising wild Vigna species also elucidated the role of proteinaceous inhibitors in imparting resistance to C. maculatus. Most of the workers attribute these inhibitors as proteins with a molecular weight of 14 to 29 kDa. In the present studies too, banding patterns were observed in the regions between 14 and 29 kDa which might be corresponding to these protease inhibitors. Oliveira et al. (1999) observed the presence of trypsin inhibitors with a molecular weight of 28 kDa that were responsible for bruchid resistance in Canavalia ensiformis. The bands corresponding to 14 kDa in the present findings may represent some form of cysteine protease inhibitors as reported by Brzin et al. (1998). They also observed a banding pattern for cysteine protease inhibitors at 27 kDa and attributed this subunit to a dimer of cysteine protease inhibitor, which was more stable than the monomer (14 kDa). Thus the intense banding at 27 kDa in the present study may correspond to cysteine protease inhibitor. This is further confirmed by the absence of the above-mentioned bands in CO 6.
Thus the present studies provide clear cut evidence on the role of trypsin and cysteine protease inhibitors to a greater extent along with chymotrypsin inhibitors in conferring resistance in V umbellata to C. maculatus. The search for genes from the wild is gaining momentum in the recent years as there is a possibility of incorporating such genes into cultivated varieties to make the latter resistant to crop pests. Thus, the present study emphasize the availability of resistant genes in the wild Vigna species which can be used in conventional breeding programmes as well as molecular approaches to evolve resistant cultivars of crop plants.
ACKNOWLEDGMENTS
We thank Dr. R. Samiyappan, Professor, Plant Pathology, for providing facilities at his lab for this study. The Department of Pulses, Tamil Nadu Agricultural University is also acknowledged for providing materials needed for the study.
REFERENCES
- Brzin, J., T. Popovic, A. Ritonja, V. Puizdar and M. Kidric, 1998. Related cystatin inhibitors from leaf and from seed of Phaseolus vulgaris L. Plant Sci., 138: 17-26.
Direct Link - Dongre, T.K., S.E. Pawar, R.G. Thakare and M.R. Harwalkar, 1996. Identification of resistant sources to cowpea weevil (C. maculatus (F.)) in Vigna spp. and inheritance of their resistance in black gram (Vigna mungo var. mungo). J. Stored Prod. Res., 32: 201-204.
Direct Link - Hines, M.E., C.I. Osuala and S.S. Nielsen, 1991. Isolation and partial characterization of soybean cystatin cysteine proteinase inhibitor of coleopteran digestive proteolytic activity. J. Agric. Food Chem., 39: 1515-1520.
Direct Link - Ignacimuthu, S., S. Janarthanan and B. Balachandran, 2000. Chemical basis of resistance in pulses to Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). J. Stored Prod. Res., 36: 89-99.
CrossRefDirect Link - Janzen, D.H., H.B. Juster and E.A. Bell, 1977. Toxicity of secondary compounds to the seed eating larvae of the bruchid beetle Callosobruchus maculatus. Phytochemistry, 16: 223-227.
CrossRef - Koiwa, H., R.E. Shade, K. Zhu-Salzman, L. Subramanian and L.L. Murdock et al., 1998. Phage display selection can differentiate insecticidal activity of soybean cystatins. Plant J., 14: 371-379.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lale, N.E.S. and M.S. Makoshi, 2000. Role of chemical characteristics of the seed coat in the resistance of selected cowpea varieties to C. maculatus (F.) (Coleoptera: Bruchidae) in Nigeria. Int. J. Pest Mgt., 46: 97-102.
Direct Link - Marconi, E., S. Ruggerei and E. Carnovale, 1997. Chemical evaluation of wild under-exploited Vigna sp. seeds. Food Chem., 59: 203-212.
Direct Link - Oliveira, A.E.A., M.P. Sales, O.L.T. Machado, K.V.S. Fernandes and J. Xavier-Filho, 1999. The toxicity of the jack bean [C. ensiformis (L.) DC] cotyledonary and testa proteins to the cowpea weevil [Callosobruchus maculatus (F)]. Entomol. Exp. Applied, 92: 249-255.
Direct Link - Saikia, P., C.R. Sarkar and I. Borua, 1999. Chemical composition, antinutritional factors and effect of cooking on nutritional quality of rice bean [Vigna umbellata (Thunb: Ohwi and Ohashi)]. Food Chem., 67: 347-352.
Direct Link - Sarikarin, N., P. Srinives, R. Kaveeta and P. Saksoong, 1999. Effect of seed texture layer on bruchid infestation in mungbean Vigna radiata (L.) Wilczek. Sci. Asia, 25: 203-206.
Direct Link - Singh, U. and B. Singh, 1992. Tropical grain legumes as important human foods. Econ. Bot., 46: 310-321.
CrossRefDirect Link