
H. Nook Farikhah
Department of Silvieulture, Faculty of Forestry, Bogor Agricultural University, P.O. Box 168, Bogor, 16680, Indonesia
A.S. Sajap
Faculty of Forestry, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
A.B. Idris Ghani
Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 4 | Page No.: 346-349







ABSTRACT
A study on the abundance lacewing Glenochrysa sp. (Neuroptera:Chrysopidae) in a lowland dipterocarp forest at different stages of recovery after longing was carried out. The lacewings were sampled using sticky traps baited with eugenol. The traps were placed for one week at a three month interval over one year in a primary forest and forests of five and ten years after logging. The result shows that the abundance of Glenochrysa sp. varied with the stages of forest recovery after logging. The number Glenochrysa sp. was significantly higher in the logged over forests than in the primary forest. Even though the lacewing was present all year round, its abundance, however, was influenced by the composition of vegetation within the selected forests, availability of food resources as indicated homopteran abundance and amount of rainfall.
PDF Abstract XML References Citation
How to cite this article
H. Nook Farikhah, A.S. Sajap and A.B. Idris Ghani, 2007. Abundance of Lacewing, Glenochrysa sp. (Neuroptera: Chrysopidae) in Forest at Various Stages of Recovery after Logging at Sungai Lalang Forest Reserve, Selangor, Malaysia. Journal of Entomology, 4: 346-349.
DOI: 10.3923/je.2007.346.349
URL: https://scialert.net/abstract/?doi=je.2007.346.349
DOI: 10.3923/je.2007.346.349
URL: https://scialert.net/abstract/?doi=je.2007.346.349
INTRODUCTION
Forest supports the greatest abundance of life forms including insects. The forest provide habitats for thousands insect species including those that serve as parasitoids and predators. These parasitoids and predators maintain the insect diversity by regulating the population of insect community in the forest ecosystem (La Salle and Gauld, 1993). Even though, parasitic Hymenoptera, predatory ants and beetles were the most commonly studied natural enemies in the tropical rain forests (La Salle and Gauld, 1993; Idris, 2000) there are also other predacious insects that concomitantly play regulatory roles in these complex forest ecosystems. One of these predacious insects, is the green lacewing represented by a Glenochrysa sp. (Neuroptera: Chrysopidae). Unlike other Malaysian species of Chrysopidae, such as Malada basalis, Ankylopteryx trimaculata and Italochrysa aequalis that are common in the agro-ecosystems, this little known lacewing, Glenochrysa sp. inhabits the tropical rain forest ecosystem. The adults predominantly occupy a stratum between 5 to 10 m above ground below the forest canopy (Sajap et al., 1997). Apart from that study, heretofore, no information related to its biology and ecology was known.
New (2002) indicates that Glenochrysa sp. is probably endemic to Malesia region and Northern Australia and several species have yet to be described. Even though many species have yet to be described, their diversity and abundance of chrysopids in the region are currently under threat for the reason that much of the Malesian region has been subject to massive environmental changes over the last century. Consequently many of these chrysopids may have probably become less diverse and less abundant. This consequent has been shown to occur on other insects but the resulting impact may vary with the insect species. For examples, a significant loss of moth diversity and taxonomic quality occurred when natural forest was converted to plantation (Holloway et al., 1992), while the beetles were less sensitive to the changes (Chung et al., (2000). The abundance of ichneumonid wasps, represented by a diverse group of species, was found to be significantly different among different habitats (Idris, 2000; Idris and Hanidah, 2003). Their numbers were invariably higher in the primary forest reserve than in other habitat types. This study examined the population abundance and fluctuation of the lacewing in forest habitats at different stages of recovery after logging.
MATERIALS AND METHODS
Study Site
The study was carried out at Sungai Lalang Forest Reserve, Semenyih, Selangor, about 50 km south of Kuala Lumpur. The forest, which covers an area of 729 hectares with an elevation of about 150-800 m above sea level, is characterized as lowland and hill Dipterocarp forest that is dominated by Shorea species.
Three compartments within this forest reserve, Compartment 33, Compartment 18 and Compartment 24, were selected for the study. Compartment 33 has an area of 387 ha and was logged 10 years ago while compartment 18 has an area of 260 ha and was logged 5 years ago. In contrast, the compartment 24 is a primary forest, which can be characterized as hill forest covering an area of 84 ha.
Lacewings sampling
Sampling was for the lacewings commenced in 1999. Prior to sampling, three plots about 500 m apart, were established along a transect line in each of the three forest compartments. The transect cut across the forest from the edge into the forest. Sticky traps made of a square nylon mesh measuring, 30x30 cm, coated with insect trapping adhesive, TanglefootR (The Tanglefoot Co. Grand Rapids, Michigan) with a 5 mL glass vial containing eugenol plugged with a cotton wick, were used for trapping the lacewings (Sajap et al., 1997). These traps, hung at 3 m height and spaced every 5 m remained in each locality for one week. The number of adult lacewings captured on the traps was counted there after. Trappings were carried out four times at three-month interval in a period of one year. A total of five traps were used in each plot.
Vegetation sampling
Nine habitat plots for vegetation sampling were established in each of the three compartments. Each plot was about 250 m2 and was further divided into 25 m2 (5x5 m) sub-plots. These plots were located in the vicinity of the lacewing sampling plot. The number of small shrubs (height <2 m), tall shrubs (height>2 m), short woody plant (height<1 m), short herbs (height<1 m), small stems (height<1 m) and stems (height 1-5 m) were recorded. All plants bellow 5 m height were considered as vegetation. The number of trees (≥5 cm dbh) was sampled from nine hectares plots that established in three forest habitats.
Homopteran sampling
In this study Homoptera was used as an indicator of food resource for the lacewings. Homoptera had been cited as the preferred hosts for many predacious chrysopid species (Canard et al., 1984). Nine malaise traps were placed in each forest compartment in the same duration with sticky trap. Content of the traps was emptied and all small homopterans were recorded.
Rainfall recording
The rainfall data for duration of the study was taken from Semenyih Dam Station.
Data collection and analysis
An analysis of variance (ANOVA) was done to examine the significant differences in the number of insect, microhabitat and microclimate attributes among the different forest habitats. Tukey’s Studentized Range (HSD) was used to compare the respective means. The relationship between the rainfall and abundance of lacewings was examined using linear regression. All statistical analysis was done using Statistical Analysis System (SAS) Program, version 6.12.
RESULTS
The trappings gave total catches of 513 adult lacewings from 5-year old logged, 406 from 10 year old logged and 239 from primary forests. Statistically, there was no significant difference between the five-year old logged forest and ten-year old logged forest but there was a significant difference in the number of individuals between primary and logged forest sites (Table 1). The number of individuals recorded from all plots within each compartment was not significantly different (Table 2).
Even though the lacewings were present all year round but their numbers fluctuated over time. Highest number of lacewings, 463 individuals, was recorded in December and they declined to their lowest, 173 individuals in June (Table 3). There was a significant relationship between number of Glenochrysa sp. and total rainfall and produced equation Y = 1.12 X - 14.69 (R2= 0.75, p<0.05) (Table 3). In addition to rainfall, the abundance of Glenochrysa sp. was positively correlated to the total number of Homoptera as its prey (r = 0.96, p<0.05) and the total number of vegetation taxa (r = 0.95, p<0.05) in the study area (Table 4). The relationship between the number of Glenochrysa sp. with the number of trees, however, was inversely correlated (r = -0.86, p<0.05) (Table 4).
| Table 1: | The mean total number of Glenochrysa sp. adult caught in the sticky trap |
 | |
| The different letter (s) are significantly different at p<0.05 | |
| Table 2: | Mean number of individuals of Glenochrysa sp. caught in the sticky traps |
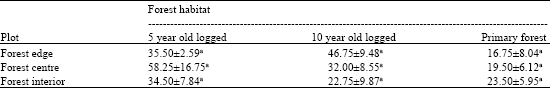 | |
| Difference letter (s) indicate significant (p<0.05) differences based on the Tukey-type multiple comparison | |
| Table 3: | The amount of rainfall (mm) and number of Glenochrysa sp. within one year |
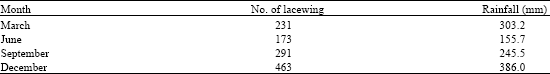 | |
| Table 4: | Total number of Glenochrysa sp. adult, homoptera, tree and understorey plant sampled in three different forest habitats |
 | |
| a: Sampled using malaise trap; b: In 1 ha plot; c: In 250 m2 plot | |
DISCUSSION
The abundance of predatory insects depends on the primarily on the availability of their prey. This prey-predator relationship was exemplified by the tropical rainforest chrysopid, Glenochrysa sp. and its homopteran prey. The abundance of prey was one of the factors that contributed to the higher number of chrysopid in the logged over forest than in the primary forest. The condition of the logged over forests with dense shrubs and trees apparently served a better habitat for the prey and its predator than the primary forest. These trees and shrubs invariably flushed during rainy seasons. During flushing, these plants supported higher number of insects, particularly sap-sucking insects, that served as preys for the chrysopid community (Wolda, 1978). Sap-sucking insects as suggested by Janzen (1973) were more sensitive to changes in the availability of young leaves and shoots than folivorous insects. Because of the larvae and most adult Chrysopidae are predaceous insects, their feeding relations to plants are indirect and they are not necessarily confined to particular plant species for their feeding requirements (Canard et al. 1984). Population of lacewing as a predator had been demonstrated to follow population of its prey, Homoptera. As prey population increase, predator population increase and as prey population decrease, predator population decrease (Price, 1997). The flight behaviour of Glenochrysa sp may also be a factor that contributed to its abundance in the logged over forests. The highest flight activity of lacewings was reported to be below 30 m (Canard et al., 1984) and that Glenochrysa species was mostly caught at 5-10 m above ground. The adult lacewings forage under the forest canopy consuming small soft-bodied arthropods in addition to honey dew, nectar and pollens.
REFERENCES
- Idris, A.B. and J. Hainidah, 2003. Diversity of ichneumonid wasps in the logged over forests of Langat basin in Selangor, Malaysia. J. Biological Sci., 3: 259-270.
CrossRefDirect Link