
K. Sahayaraj
Department of Advanced Zoology and Biotechnology, Crop Protection Research Centre, St. Xavier�s College (Autonomous), Palayamkottai-627002, Tamil Nadu, India
S. Kumara Sankaralinkam
Department of Advanced Zoology and Biotechnology, Crop Protection Research Centre, St. Xavier�s College (Autonomous), Palayamkottai-627002, Tamil Nadu, India
R. Balasubramanian
Department of Advanced Zoology and Biotechnology, Crop Protection Research Centre, St. Xavier�s College (Autonomous), Palayamkottai-627002, Tamil Nadu, India
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 4 | Page No.: 331-336







ABSTRACT
Rhynocoris marginatus (Fab.) was reared on two natural lepidopteron pests [Spodoptera litura (Fabricius) and Pericallia ricini (Fab.)] and a factitious host (Corcyra cephalonica Stainton). Whereas Catamiarus brevipennis (Serville) reared on C. cephalonica, S. litura and Thunberg. Impact of these preys on the total body, midgut, hindgut and entire salivary gland weight, their enzymes qualitative profile was recorded. Total carbohydrates and lipids and more amount of proteins were observed in S. litura fed R. marginatus showed maximum total body, midgut, hindgut and salivary gland weights. Presence of more amount of total lipids and carbohydrates in M. pustulata favours the body, gut and salivary gland weight of C. brevipennis. Amylase, invertase, lipase, protease, trypsin and pepsin activities were well pronounced in the midgut than the hindgut of both reduviids. R. marginatus and C. brevipennis salivary gland expressed more amylase, invertase, protease and lipase activities.
PDF Abstract XML References Citation
How to cite this article
K. Sahayaraj, S. Kumara Sankaralinkam and R. Balasubramanian, 2007. Prey Influence on the Salivary Gland and Gut Enzymes Qualitative Profile of Rhynocoris marginatus (Fab.) And Catamiarus brevipennis (Serville) (Heteropetera: Reduviidae). Journal of Entomology, 4: 331-336.
DOI: 10.3923/je.2007.331.336
URL: https://scialert.net/abstract/?doi=je.2007.331.336
DOI: 10.3923/je.2007.331.336
URL: https://scialert.net/abstract/?doi=je.2007.331.336
INTRODUCTION
Rhynocoris marginatus (Fab.) native to India is found to feed on more than 24 economically important pests including Earias fraterna (Pawar et al., 1986; Ambrose, 1988), Dysdercus cingulatus (Fab.) (Imms, 1965), Papilio demoleus (L), Earias vittella (Fab.) (Nayer et al., 1976); Corcyra cephalonica (Stainton) (Bhatnagar et al., 1983); Helicoverpa armigera (Hubner) (Ambrose, 1987); Spodoptera litura (Fab.), Amsacta albistrigia (Walker) (Sahayaraj, 2000); Mylabris pustulata (Faust), Mylabris indica (Thunberg), Achaea janata (Linn.), Oxycarenus hyalinipennis (Costa) and Aproarema modicella (Deventer) (Sahayaraj, 1995; Sahayaraj and Kathikraja, 2003). Pest suppression efficacy of this predator was studied both under laboratory (Imms, 1965; Pawar et al., 1986) and field conditions (Sahayaraj, 1999; Sahayaraj and Martin, 2003) in India. Scope for the utilisation of R. marginatus in groundnut pest management was emphasised by Sahayaraj (1999, 2004) and Sahayaraj and Martin (2003).
Catamiarus brevipennis (Serville) is one of the larger predator of the family Reduviidae and sub-family Peiratinae present in scrub jungles, semi-arid zone, tropical rain forest and agroecosystems of south India (Sahayaraj, 1994). It has been reported as a biological control agent of many agriculture and forest pest like H. armigera (Bhatnagar et al., 1983), Pantanga succincta (Linn.) (Pawar et al., 1986), Earias insulana (Fab.) and Mylabris pustulata Thunberg (Ambrose, 1987), D. cingulatus and Achaea janata (Linn.) (Sahayaraj, 1991). Sahayaraj (1991) and Sahayaraj and Ambrose (1994) studied the host preference of this reduviid on various pests. However, works on biology, ecology and biological control potential of this reduviid has not been undertaken either under laboratory or field situations.
The digestive physiology of reduviid predators solicits greater attention in view of its economic importance. The nutritional need and the knowledge of the functional organisation of digestive system of reduviid predators may be useful in designing oligidic diet for mass production. Moreover this information can be useful to understand how reduviids adopt to its natural and factitious food. Enzymes are proteins, which catalyse a variety of reactions in the biological systems. Digestive enzymes are produced and distributed in different regions of the gut and differ in proportion and quantity. Wide ranges of digestive enzymes were recorded in the alimentary canal of insects and the level varies in relation to moulting and short-term changes in food intake. Food stimulates the secretion of protease in the midgut, the intake of insect substances like water and cellulose. It is a well-known fact that the digestive enzymes play a major role in insect physiology by converting complex food materials into micromolecules necessary to provide energy and metabolites for growth, development and other vital functions. Among the digestive enzymes, protease, amylase, invertase and lipase activity are of great importance in the digestion of food. Utilization of macronutrients from the available prey food depends on the digestive enzymes. Digestive enzymes of alimentary canal and the salivary gland of Sophrorhinus insperatus Faust (Hori, 1969; Ravikumar et al., 2002) were investigated. Influence of host plants on the activity of digestive enzymes of Helicoverpa armigera (Hubner) was also studied. No concrete work has been undertaken so far about digestive enzymes in predaceous reduviids. Digestive enzymes of Heteroptera include proteinase, lipase, phospholipase A1, amylase, pectinase, invertase, hyaluronidase and nuclease (Nuorteva, 1958; Miles, 1972; Cohen, 1998). Digestive enzymes are specific for zoophagic insects include protease, hyaluronidase and phospholipase (Cohen, 1998). However studies on enzyme profile of Indian reduviids were not available in the literature. Based upon the available literature we have undertaken this research to record the qualitative enzyme profile of salivary gland, mid and hindgut of R. marginatus and C. brevipennis.
MATERIALS AND METHODS
Collection and Maintenance of Reduviids
Life stages of R. marginatus and C. brevipennis were collected from Sivanthipatti agricultural ecosystem (cotton and bhendi) and also their border ecosystems of scrub jungles, Palayamkottai, Tirunelveli District, Tamil Nadu, India. They were maintained in the laboratory at 28±1°C temperature, 70±10% relative humidity and 11 L: 13 D on C. cephalonica, M. pustulata, P. ricini and S. litura.
Rearing of Preys
Larval stages of S. litura, M. pustulata and P. ricini were collected from cotton and ladyfinger fields in Pavoorchatram, Tirunelveli District, Tamil Nadu, India and were maintained in the same laboratory conditions as for reduviids on castor and black gram leaves. Laboratory emerged fifth instars S. litura, P. ricini and field collected M. pustulata adults were used for the experiments. Newly emerged adult of R. marginatus were reared on S. litura, P. ricini and C. cephalonica whereas C. brevipennis reared on C. cephalonica, S. litura and M. pustulata continuously for three weeks separately. Then the predators were removed from the host and allowed to starve for 6 hours. Twenty predators from each category were used for the enzyme profile analyses studies. Ten uniform sized preys were selected for the macromolecules profile. Total carbohydrates (Nigam and Omkar, 2003), total proteins (Lowry et al., 1951) and total lipids (Bragdon, 1951) were estimated with standard procedures.
Enzyme Extraction
Both the reduviid adults (>24 h) were maintained on S. litura and C. cephalonica continuously for a period of three weeks separately. Anesthetized predators were dissected out aseptically by using pins, fine forceps and razors and a dissection microscope in a petri dish filled with sterile phosphate buffered saline (pH 7.8). Entire gut and salivary gland were dissected out from the predators. From the isolated digestive tract, mid and hind guts were separated individually, washed several times with fresh phosphate buffered saline to minimize possible microbial contamination and used for enzyme bioassay. Similarly entire salivary gland from 10 predators were removed carefully and used for the enzyme study. Transfer the salivary glands and alimentary canal separately in a small test tube containing 2 mL of distilled water and grind them as completely as possible with tissue Homogenizer (Remi 8000 RPM, Mumbai). Transfer the solution to the centrifuge tube and centrifuge at 5000 rpm for 15 min. The supernatant was used as enzyme source for this experiment. Invertase, amylase, lipase (Nigam and Omkar, 2003), pepsin, polypeptidase (Tonapi, 1996), protease, trypsin (Balagun and Fisher, 1970), qualitative profiles were performed from the enzyme samples using sugar, starch, olive oil emulsion, acid casein, peptone, casein, alkaline casein, respectively as substrates. Based upon the colour intensity, the enzyme activities were expressed as less (+), moderate (++) and maximum (+++) activities.
Statistical Analysis
Both for macromolecules and weight of different parts of the reduviids, student t-test was performed. Results of C. cephalonica were compared with other pests and their significance was expressed at 5% level.
RESULTS
Digestive enzymes play a major role by converting complex food in to the micromolecules which are necessary to provide energy and metabolites (Wigglesworth, 1972) to the insects. Macromolecules weight of tested preys is presented in Table 1. From the results, it was very clear that total carbohydrates, total proteins and total lipid contents were higher in C. cephalonica, S. litura and M. pustulata, respectively.
Gut Enzyme Profiles
The food quality regulates and influences the production of digestive enzymes. Henceforth, an attempt was made to study the qualitative profile of digestive enzymes in relation to different preys. From the Table 2, it is very clear that both the reduviids midgut and hindgut contain amylase, protease, invertase, lipase, trypsin and pepsin. But their activity profile is depends on the type of host encountered. For instance, protease and lipase activities of both the mid and hindgut were maximum while R. marginatus was provided on C. cephalonica. But both the trypsin and pepsin levels of midgut were higher (+++) when the predator consumed S. litura. C. brevipennis midgut amylase, protease, invertase, lipase, trypsin and pepsin activities were maximum same on the other preys except on M. pustulata (Table 2). However, the hindgut enzyme activities were decreased (++) and similar kind of observations were recorded for all the three preys. The results further showed that prey type does not have any influence on the enzyme activity of this reduviid.
| Table 1: | Macromolecule composition (mg/mL) of four pests (mg/100 mg) |
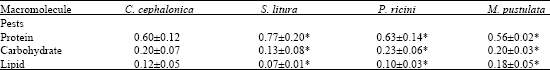 | |
*: Significant at 5% level | |
| Table 2: | C. cephalonica (CC), S. litura (SL), M. pustulata (MP) and P. ricini (PR) influence on the midgut and hindgut qualitative enzyme profile of R. marginatus |
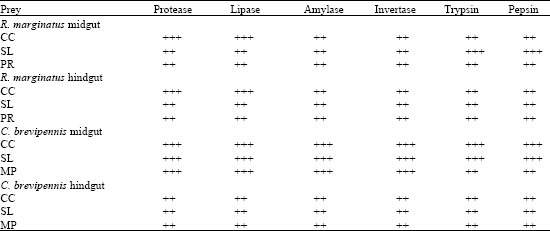 | |
| (++) Moderate; (+++) Maximum | |
| Table 3: | Influence of C. cephalonica (CC), S. litura (SL) and M. pustulata (MP) on salivary gland enzyme profile of R. marginatus and C. brevipennis |
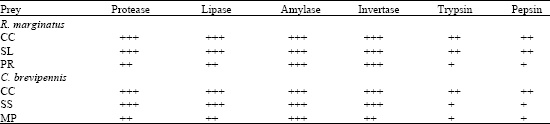 | |
(+) Less, (++) Moderate, (+++) Maximum | |
| Table 4: | Influence of preys on body, gut and salivary gland weight (mg) of C. brevipennis and R. marginatus |
 | |
| * Shows significant at 5% level by t-test; NS- Stars for not significant | |
Salivary Gland Enzyme Profile
Polypeptidase activity was not observed in the salivary gland of both R. marginatus and C. brevipennis (Table 3). Both the trypsin and pepsin activities were moderate (++) when R. marginatus was provided with C. cephalonica and S. litura and C. brevipennis on C. cephalonica. The activity was further reduced (+) on P. ricini for R. marginatus, S. litura and M. pustulata for R. marginatus and C. brevipennis, respectively. In R. marginatus and C. brevipennis, the amylase, invertase, lipase and protease activities were higher with C. cephalonica and S. litura.
Body and Body Parts Weight
Table 4 showed the total body, alimentary canal and salivary glands weight of R. marginatus and C. brevipennis. The weight was varied while R. marginatus provided with different types of prey. Statistically significant low body (129.94±2.14 mg), midgut (7.54±1.21 mg), hindgut (6.98±1.10 mg) and salivary glands were recorded while R. marginatus was fed with P. ricini. From the Table 4, it was very clear that S. litura was the suitable prey for the rearing of R. marginatus. But the statistical comparison between S. litura and C. cephalonica were insignificant. Body weight was higher while R. marginatus was reared on S. litura (134.7 mg). But it was not statistically insignificant when compared to P. ricini and C. cephalonica. Similar statistical insignificance was also recorded for salivary gland weight (Table 4). M. pustulata slightly influence C. brevipennis weight (779.33 mg). It was statistically significant at 5% level. However, prey has no influence on salivary gland (64.40, 65.45 and 65.78 mg for C. cephalonica, S. litura and 65.78 mg, respectively) as well as alimentary canal weight (Table 4). Body weight was statistically increased when C. brevipennis provided with both S. litura and M. pustulata. Similar trend was also observed for midgut.
DISCUSSION
All the hunter reduviid bugs are insect-feeders and this ability requires them to have a good complement of digestive enzymes so as to bring about hydrolysis of the complex nutrients that characterize insects. Reduviids inject toxic salivary secretion in to the host during paralysing act and consumed the body content of the victim (Sahayaraj, 2004). Cohen (1998) reported that reduviid consume partially digested food. Before consuming the host, predators predigest the host with the help of the salivary enzyme. Then ingest the partially digest proteins, carbohydrates and lipids and other nutrients, which can be further digested with alimentary canal enzymes. As observed in other insects, the production of midgut enzymes is not a continuous process and the level varies in relation to moulting and food intake (House 1905; Isaiarasu et al., 2003). Furthermore Khan (1964) in Locusta migratoria and Ishaaya et al. (1971) in S. litura, the secretion of digestive enzyme were stimulated by the intake of food. Digestive enzymes are vital determinants for growth and survival of predatory insects.
ACKNOWLEDGMENTS
Senior author thank the CSIR (ref. No. 37 (1166)/03/EMRII/), Government of India, New Delhi for the financial assistance. Our sincere thanks also to Rev. Dr. A. Antonysamy S.J. Principal and Prof. M. Thomas Punithan, Head, Department of Advanced Zoology and Biotechnology, St. Xavier’s College, Palayamkottai for the laboratory facilities.
REFERENCES
- Balogun, R.A. and O. Fisher, 1970. Studies on the digestive enzymes of the common African toad Bufo regularis boulenger. Comp. Biochem. Physiol., 33: 813-820.
CrossRefDirect Link - Bhatnagar, V.S., S. Sitanathan, C.S. Pawer, D. Jadhav, V.R. Rao and W. Reed, 1983. Conservation and augmentation of natural enemies with reference to integrated pest management in chickpea (Cicer arietinum L.) and pigeonpea (Cajanus cajan L.) Mill sp. Proceedings of the International Workshop on Intergrated Pest Control for Grain Legumer-Goiania Oias, April 4-9, 1983, Brazil, pp: 57-180.
- Hori, K., 1969. Effect of various activators on the salivary amylase of the bug Lugus disponsi. J. Insect. Physiol., 15: 2305-2317.
CrossRef - Isaiarasu, L., M. Krishnan and S. Mathavan, 2003. Effect of dietary glycine on the activity of some digestive enzymes in the final instar larvae of Bombyx mori L. Entomon, 28: 71-76.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link