
P. Balaji
Lichen Ecology and Bioprospecting Laboratory, M.S. Swaminathan Research Founthtion, Ill Cross Street, Taramani Institutional Area, Chennai 1 3, India
S. Malarvannan
Biopesticides Laboratory, MSSRF, India
G.N. Hariharan
Lichen Ecology and Bioprospecting Laboratory, M.S. Swaminathan Research Founthtion, Ill Cross Street, Taramani Institutional Area, Chennai 1 3, India
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 3 | Page No.: 248-252







ABSTRACT
The insecticidal activities of hexane, ethyl acetate, methanol and water extracts of lichen, Roccella montagnei Bél. emend. Awas. were assayed against polyphagous insect, Helicoverpa armigera (Hübner). Among the different crude extracts tested, the pupation was nil in methanol (1%) and hexane extract (5%), which was on par with aqueous extract (5%). The formulation of water extract of R. montagnei is better compared to other solvent extracts, as potential antipest agent against H. armigera.
PDF Abstract XML References Citation
How to cite this article
P. Balaji, S. Malarvannan and G.N. Hariharan, 2007. Efficacy of Roccella montagnei Extracts on Helicoverpa armigera (Lepidoptera: Noctuidae). Journal of Entomology, 4: 248-252.
DOI: 10.3923/je.2007.248.252
URL: https://scialert.net/abstract/?doi=je.2007.248.252
DOI: 10.3923/je.2007.248.252
URL: https://scialert.net/abstract/?doi=je.2007.248.252
INTRODUCTION
Gram caterpillar, Helicoverpa (= Heliothis) armigera (Hübner) (Lepidoptera: Noctuidae) causes heavy infestation in more than sixty-seven cultivated species. It has higher mobility and fecundity (Reed and Pawar, 1982) and higher resistance to chemical pesticides. This has led to the usage of extracts of Azadirachta indica, Vitex nigundo (available as commercial products), Ocimum sanctum, Tagetes erecta, Tribulus terrestris, Capsicum annum, Cipadessa baccifera, Clausena dentata, Dodonaea angustifolia and Melia dubia (Malarvannan, 2004). The above mentioned botanicals are preferred to synthetics due to their multiple action on the insect’s life cycle stages, low cost, easy availability and least toxic to environment (Kumar and Sangappa, 1984; Bruno et al., 1999). Similarly, the unique secondary compounds of lichens such as aliphatic acids, pulvinic derivatives, hydroxybenzoic acids, depsides, depsidones, dibenzofurans, anthroquinones, naphthaquinones are also known to exert antiherbivore, antibiotic and other effects (Huneck, 1999; Müller, 2001; Rundel, 1978; Lawrey, 1984, 1989). Even though these manifold activities of lichen metabolites have now been recognized, their potentials have not yet been fully explored and thus remain unexploited (Müller, 2001). This paper reports the efficacy of the organic solvent extracts of Roccella montagnei Bél. emend. Awas. (Ascomycotina) on H. armigera.
MATERIALS AND METHODS
The thalli of R. montagnei were collected from Rhizophora apiculata trees from Pichavaram (11° 23′ N to 11° 30′ N latitude and 79° 45′ E to 79° 50′ E longitude). Voucher specimens (MSSRF 207/02, 208/02) and deposited at the Lichen Ecology and Bioprospecting Laboratory, M.S. Swaminathan Research Foundation, Chennai, India. The thalli of R. montagnei (950 g) were shade dried at room temperature and serially extracted with hexane, ethyl acetate, methanol and water (5 L) successively by Soxhlet extractor.
| Table 1: | Effect of R. montagnei extracts on growth inhibition of H. armigera |
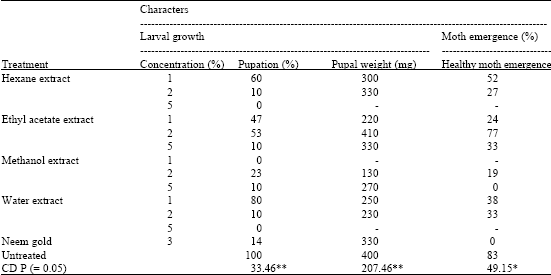 | |
| *Each value a mean of triplicate | |
The pooled solution was evaporated using a rotary evaporator (Buchi rotavapor R-200) to obtain the crude extract. The crude extract was lyophilized to powder (VirTis bench top model) and stored at -30°C.
The larvae of H. armigera collected from lady’s finger, Abelmoschus esculentus were reared in the laboratory on semi-synthetic diet (Shorey and Hale, 1965) in individual containers to prevent cannibalism and contamination and maintained at 22±2°C and 70-75% RH. The experiment was conducted at M. S. Swaminathan Research Foundation, Chennai during 2005-06. Healthy third instar larva was chosen for the experimentation. Crude hexane, ethyl acetate, methanol and water extracts (1, 2 and 5%) of R. montagnei were mixed with semi-synthetic diet. Neem gold (positive control) and untreated diet (normal diet-negative control) were also maintained for comparison. The larvae were released into the treated diet after 1 h of starvation. Five replications with 10 larvae per replication was maintained and the parameters such as pupation, pupal weight and healthy moth emergence (Table 1) were critically monitored and recorded. The data were statistically analyzed using Agres package (Panse and Sukhatme, 1967) to understand the critical difference between the performance of various treatments using single ANOVA.
RESULTS AND DISCUSSION
At 1% level extract treatment, the percentage of pupation was nil in larvae treated with methanol extract and 80% in aqueous extract. At 2% level, 10% pupation was observed in both hexane and aqueous extracts and 53% in ethyl acetate extract. At 5% level, no pupation was observed in hexane and aqueous extract while 10% pupation was observed in larvae treated with both ethyl acetate and methanol extracts of R. montagnei.
Among the different extracts tested at 1% level, the pupal weight of ethyl acetate extract treated larvae was 220 mg while it was 300 mg in hexane extract and 400 mg in the untreated (control). At 2% level, 130 mg of pupal weight was observed in methanol and 410 mg in ethyl acetate. At 5% level, the pupa weighed 270 mg in methanol and 330 mg in ethyl acetate.
The healthy moth emergence of H. armigera at 1% level of ethyl acetate extract treatment was 24 and 52% in hexane extract. At 2% level, 33% of moth emergence was observed in aqueous extract and 77% in ethyl acetate extract. At 5% level nil moth emergence was observed in methanol while it was 33% in ethyl acetate. In control, the healthy moth emergence was 84%.
In addition to larval mortality, 1% level of hexane, methanol and aqueous extracts showed malformed and shriveled larvae. At 2% level aqueous extract treated larvae showed delayed moulting. At 5% level, disturbed moulting, delayed pupation (up to 10-12 days) and malformations were high irrespective of the extracts tested. Moreover, pupation in hexane (2%) and aqueous (5%) extract was delayed and resulted in discoloration (pale yellow) and drastic reduction in their weight. Contrastingly, pupal weight was high (410 mg) in ethyl acetate, which was more than untreated (400 mg). Regarding moth emergence, the adults emerged from hexane, ethyl acetate and methanol extracts had irregular wing size and loss of appendages.
In our experiments, no pupation was observed in methanol (1%), hexane (5%) and aqueous extract (5%) of R. montagnei. Among those pupated, it was least (10%) in hexane and water extract at 2% as well as ethyl acetate and methanol at 5% concentration (Table 1). Lawrey (1984) observed nil effect on larval growth or survival due to lichen compounds (Cetraria pinastri) against Spodoptera ornithogalli. The (-) and (+)-usnic acid and vulpinic acid, 3-hydroxyphysodic provoked strong mortality as well as significant growth retardation in Spodoptera littoralis (Emmerich et al.,1993; Giez et al., 1994). Other than insects, such reduction in feeding and subsequent growth disturbances was observed due to lichens, Letharia vulpina against slug, Deroceras reticulatum (Lawrey, 1983, 1989) and caperatic acid (Lawrey, 1983, 1989) and extracts of Flavoparmelia baltimorensis and Xanthoparmelia cumberlandia have antiherbivore activities against the snail Pallifera varia (Clark et al., 1999). Lichen secondary compounds Methyl β-orcinolcarboxylate, ethyl hematommate and 5-chlorohematommate showed nematicidal activity on larvae of Toxocara canis (Ahad et al., 1991).
The larval, pupal and moth emergence in H. armigera is a complex process involving physical (both external for e.g. temperature and internal for e.g., Midgut pH), physiological and metabolic processes. In our experiments significant reduction in pupation with larval-pupal intermediates (due to phagodepression and difficulty in moulting) was observed in hexane, ethyl acetate and methanol extracts of R. montagnei. The decrease in the juvenile hormone titre and its associated disturbances in oogenesis, larval-pupal and pupal-adult are interpreted as an interference with moulting hormone pools (Rembold et al., 1982). Decrease in juvenile hormone influences the storage proteins and fat bodies, which are highly essential for metamorphosis, moulting and reproduction (Palli and Locke, 1987; Koul and Isman, 1991). In this, study, the nil pupation and related effects can be attributed to these concentrations that have disturbed the moulting hormone pools, which are core factors for significant juvenomimetic activity.
Pupal weight was least in 2% methanol treatment (130 mg) followed by 1% ethyl acetate (220 mg) as against 400 mg in the untreated. The healthy moth emergence was nil in 5% methanol, which was on par with the Neem gold (3%) followed by 19% in methanol extract at 2% level. The suppression/reduction in pupal growth/weight was observed in lichen extracts and this effect may be due to the inhibitory action on mitochondrial respiration by affecting the NADH- Cytochrome C-reductase and complex-I of insect mitochondria as observed in insect as well as mammalian mitochondria by Londershausen et al. (1991) and Degli Esposti et al. (1994).
Species with an evolutionary history of feeding on heavily chemically defended plant structures should have elevated levels of enzymes that detoxify defensive chemicals and therefore an enhanced ability to evolve resistance to synthetic toxins (Dominguez-Gily and McPheron, 1999). In the case of plant pests, especially H. armigera, the chemical constituents of plants are a significant part of the environment, a part that has been shown to affect the action of many resistance mechanisms. Hence it is vital to develop effective strategies to puzzle the insect resistance mechanism especially by introducing potential antipest compounds from sources unfamiliar (lichen secondary compounds) to the target insect i.e., H. armigera alternating with other pesticides. This strategy is also expected to affect the generalized reciprocal evolutionary relationship (the lines of evolutionary arms race) (Dawkins and Krebs, 1979) between the insect and its host, resulting in decreased fitness of the insect due to the failure of resistance mechanism. Such strategies may provide significant and long-lasting effect on the pest and keep them under economic injury level for contamination free environment and sustainable agriculture.
ACKNOWLEDGMENTS
The authors thank Prof. M. S. Swaminathan and the Executive Director, MSSRF for providing opportunity and facility to carry out the study and Prof. P.C. Kesavan, MSSRF for valuable suggestions and constant encouragement. We thank Dr. H.D. Subashini, Mr. R. S. Satyan, P. Bharath for their timely help and R. Kathiravan for laboratory assistance. We thank Department of Biotechnology (DBT), Government of India and CSIR, New Delhi for the financial support.
REFERENCES
- Clark, S.J., I.F. Henderson, D.J. Hill and A.P. Martin, 1999. Use of lichen secondary metabolites as antifeedants to protect higher plants from damage caused by slug feeding. Ann. Applied Biol., 134: 101-108.
Direct Link - Degli Esposti, M., A. Ghelli, M. Ratta, D. Cortest and E. Estornellt, 1994. Natural substances (acetogenins) from the family Annonaceae are powerful inhibitors of mitochondrial NADH dehydrogenase (Complex I). Biochem. J., 301: 161-167.
Direct Link - Giez, I., O.L. Lange and P. Proksch, 1994. Growth retarding activity of lichen substances against the polyphagous herbivourous insect Spodoptera littoralis. Biochem. Syst. Ecol., 22: 113-120.
Direct Link - Koul, O. and M.B. Isman, 1991. Effects of azadirachtin on the dietary utilization and development of the variegated cutworm, Peridroma saucia. J. Insect Physiol., 37: 591-598.
CrossRef - Muller, K., 2001. Pharmaceutically relevant metabolites from lichens. Applied Microbiol. Biotechnol., 56: 9-16.
CrossRefDirect Link - Rembold, H., G.K. Sharma, Czoppelt and C. Schmutterer, 1982. Azadirachtin: A potent insect growth regulator of plant origin. Zeitschrift Angewandte Entomol., 93: 12-17.
CrossRef - Shorey, H.H. and R.L. Hale, 1965. Mass rearing of the larvae of nine noctuid species on a simple artificial medium. J. Econ. Entomol., 58: 522-524.
CrossRefDirect Link