
S. O. Okoth
Trypanosomiasis Research Centre-Kenya Agricultural Research Institute, Box 362, Kikuyu Kenya
E.D. Kokwaro
School of Pure and Applied Sciences, Kenyatta University, Box 43844 Nairobi
J.M. Kiragu
Trypanosomiasis Research Centre-Kenya Agricultural Research Institute, Box 362, Kikuyu Kenya
G.A. Murilla
School of Pure and Applied Sciences, Kenyatta University, Box 43844 Nairobi
Journal of Entomology
Year: 2007 | Volume: 4 | Issue: 1 | Page No.: 20-32







ABSTRACT
Vector-host contact is a key epidemiological parameter in vector transmitted parasitic infections. The interaction between vectors and hosts is influenced by both environmental and endogenous factors that regulate their diurnal activity. In this study, effects of temperature, relative humidity and lax on the activity on tsetse flies and hosts were evaluated in two allopatric populations of G. pallidipes to estimate vector-host contact and risk of disease transmission within the dry and wet seasons. Results showed that vector activity was majorly influenced by temperature and relative humidity and the activity pattern was sensitive to seasons. Host movement significantly influenced vector movement in both dry and wet seasons at Nguruman, but only to a limited extent during the wet season at Busia. Host movement correlated positively with vector activity at high host densities while there was no definite correlation at low host densities. Vector-host contact was independent of prevailing weather conditions and seasons but was explained largely by random variability ascribable to innate spontaneous circadian rhythm of the vectors and the hosts. This implies that management of transmission risk in these two areas would largely depend on avoidance of contact between the vectors and hosts. This would require strategic livestock grazing patterns that is out-of-phase with peak vector activity periods to minimize contact.
PDF Abstract XML References Citation
How to cite this article
S. O. Okoth, E.D. Kokwaro, J.M. Kiragu and G.A. Murilla, 2007. Diurnal Activity of Allopatric Populations of Glossina pallidipes and Hosts and their Implications on Risk of Transmission of Sleeping Sickness in Kenya. Journal of Entomology, 4: 20-32.
DOI: 10.3923/je.2007.20.32
URL: https://scialert.net/abstract/?doi=je.2007.20.32
DOI: 10.3923/je.2007.20.32
URL: https://scialert.net/abstract/?doi=je.2007.20.32
INTRODUCTION
Tsetse flies show pronounced diurnal rhythm of activity, both in the laboratory and in the field and the patterns vary among the three taxonomic groups of Glossina (Buxton, 1955; Brady, 1972; Harley, 1965). The palpalis group is generally active in the middle of the day; the morsitans group is most often active in the morning and afternoon and the fusca group is often strongly crepuscular (Makumi et al., 1998). The pattern may also vary with sex, hunger, pregnancy and nutritional state of different components of the population, as well as being affected by other mediating environmental factors such as temperature, light intensity, odours, wind direction and speed, physiological state and colour (Leak, 1999). The activity pattern may also vary among allopatric populations as was reported by Van Etten, (1982). Other authors have similarly reported variations in the activity patterns among different subpopulations of G. pallidipes (Leak, 1999; Owaga et al., 1993; Van Etten, 1982; Vanderplank, 1941). Vector activity rhythms have important influences on transmission challenge posed to hosts with the greatest potential risk spanning peak activity periods (Leak, 1999). Similarly host diversity, distribution and abundance influence transmission risk to a host species since vectors select preferred hosts from the pool of available hosts (Wamwiri, 2005; Tarimo, 1980). In addition to host diversity, distribution and abundance, the level of synchrony between host and vector activity pattern is also paramount in determining source of blood meal by tsetse flies (Pollock, 1980; Leak, 1999). Contact can therefore be used as an epidemiological indicator of transmission risk. Since environmental factors are compounding elements in vector-host contact, understanding their contribution as covariate risk factors would be important while designing appropriate risk management strategies sensitive to spatial and temporal changes in weather and seasons. In this study therefore, the influence of environmental factors on vector-host interactions were evaluated and strategies for management of transmission risk through reduced vector-host contact across seasons is discussed with specific reference to human and livestock under peasantry and agro-pastoralists farming systems.
MATERIALS AND METHODS
Study Areas
This study was conducted at Busia and Nguruman between January and September 2005. Busia study area lies between latitude 0° 136’ South and 0° North and longitudes 33° 54’ east and 340 25’ 24’= East (Fig. 1). The area is infested with G. f. fuscipes along the riparian forest patches and G. pallidipes, which has patchy distribution, associated with woody hillside vegetation (Ford, 1971). Nguruman lies at latitude 1° 55’ S and longitude 35° 25’ E on the floor of the rift valley in southern Kenya (Fig. 2). The area is infested by G. pallidipes and G. longipennis within the woodlands and G. swynnertoni on the adjoining escarpments (Brightwell et al., 1997).
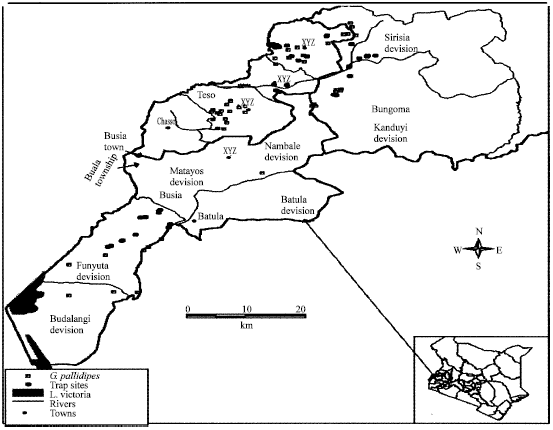 | |
| Fig. 1: | Map of Westerns Kenya showing the study area |
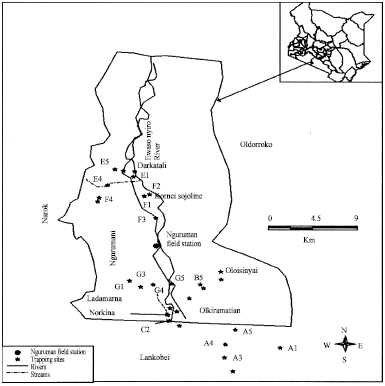 | |
| Fig. 2: | Map of Nguruman showing the study area |
Experimental Design
Two permanent trapping sites, situated at least 200 m a part, were selected in each study area on the basis of vegetation cover, proximity to human habitation and presence of hosts. At each trapping site, a single unbaited biconical trap was set for four days in February, April or August. These were to coincide with the main seasons (dry and wet) in the two study areas. The trap catches were collected at half hourly intervals from 0600-1800 h. Flies caught were sorted by sex as males and females and by age as tenerals and non-tenerals. The number of humans, livestock and wild animals sighted within 10-30 m of a trap were recorded every 10 min by an observer who approached the site from about 50 m away.
Recording of Weather Variables
Ambient temperature (°C), percent Relative Humidity (RH) and light intensity (lux) were simultaneously recorded at both trapping sites at 30 min intervals. Temperature was measured using electronic minimum and maximum thermometer (MODEL RS 212-130 China) placed under shade at a height of one meter above the ground level. The relative humidity was measured by wet-bulb whirling hygrometers and light intensity by a digital lux meter (Lutron MODEL Lx-101 Taiwan).
Data Management
The data was entered in Excel spreadsheet and analyzed using Minitab 13.0 or Statistical Package for Social Sciences (SPSS), version 9.0. Host data, light intensity, percent relative humidity and fly caches data were transformed to the logarithmic scale {log10 (n + 1)}. These were later de-transformed before interpretation. The activity patterns of male and female tsetse and of combined sexes were obtained by plotting the mean hourly catches of either sex or totals against time per season. Similar profiles were plotted for host prevalence. The peak catches of flies were identified from these plots and superimposed on those of hosts using the methods of Mohammed Ahmed and Odulaja (1997). Correlation analysis was used to determine the relationships between fly catches, weather factors and host prevalence lagged 0, 1 and 2 h to account for delayed responses. Step-wise regression analysis was used in determining key predictors in the vector activity as defined by seasonal variations and weather variables.
RESULTS
Seasonal Changes in Diel Activity Pattern of Glossina pallidipes and Hosts at Busia and Nguruman
Dry Season Activity
The activity pattern of G. pallidipes at Busia during the dry season was bimodal for both sexes. Females had morning activity peak at about 1130 h while males had slightly delayed morning activity peaking at around 1230 h. The morning peaks were equimodal for both the sexes with afternoon occurring at about 1500 h. Females were significantly more active than males during the afternoon peak as indicated by the error bars. When both sexes were combined, a unimodal diel activity profile with a slight notch between 1230 and 1400 h was obtained (Fig. 3). The morning activity begun by 0900 h reaching its peak at 1100 h, while the afternoon peak was at about 1500 h after which there was a steady decline in activity reaching zero by about 1800 h. On the other hand, the activity cycle of G. pallidipes at Nguruman showed a low morning peak and a higher evening peak with catches increasing with time into the night (Fig. 4). Morning activity of flies started as early as 0700 h with males being more active than females and peaks at 1100 h for both sexes. Males activity declines rapidly to minimum by 1300 h, while females only exhibit a slightly depressed activity that regains momentum, parallel to but higher than that of males, to highest peak at dusk. The activity profile for combined male and female G. pallidipes at Nguruman show two prominent peaks during the dry season, a morning and an afternoon peak. The activity increased gradually in the morning and peaked at 1130 h before declining slightly between 1230 and 1600 h. The evening activity was more vigorous, increasing steadily to peak at 1800 h and presumably continuing in to the night. The evening activity was also more pronounced compared to the morning fly activity.
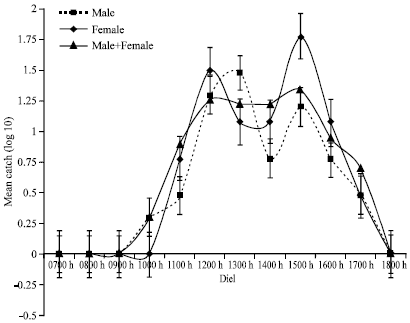 | |
| Fig. 3: | Diel activity profile for G. pallidipes at Busia during the dry season |
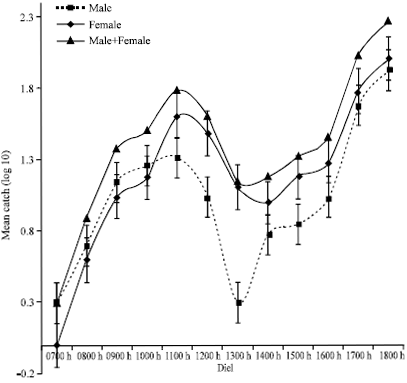 | |
| Fig. 4: | Diel activity profiles of Glossina pallidipes at Nguruman during the dry season |
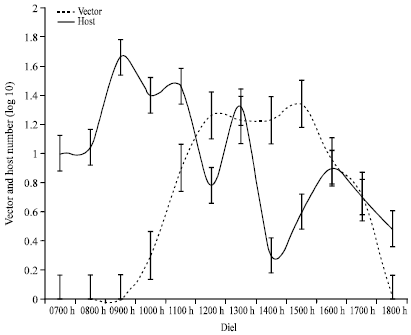 | |
| Fig. 5: | The relationship between diel activity profiles of vector and host at Busia during the dry season |
Figure 5 shows vector and host activity profiles at Busia during the dry season. It was observed that both the morning and evening peaks of the vector did not coincide with those of the host. However, there was significant interaction between the vector and the host between 1000 and 1400 h.
 | |
| Fig. 6: | The relationship between diel activity profiles of vector and host at Nguruman during the dry season |
 | |
| Fig. 7: | Diel activity profiles of G. pallidipes at Busia during the wet season |
 | |
| Fig. 8: | Diel activity profiles G. pallidipes at Nguruman during the wet season |
 | |
| Fig. 9: | The relationship between diel activity profiles of vector and host at Busia during the wet season |
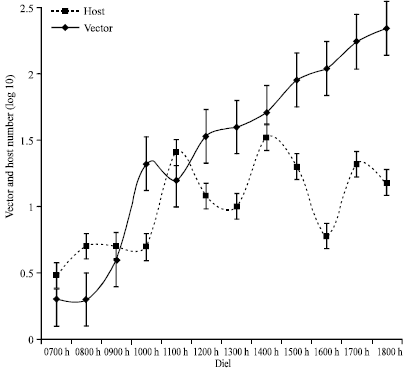 | |
| Fig. 10: | The relationship between diel activity profiles of vector and host at Nguruman during the wet season |
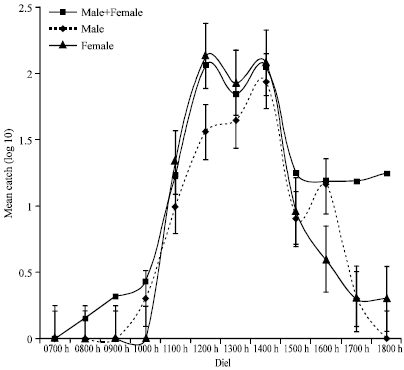
Regression analysis with vector as response and host as predictor variable defined the relationship as: Vector activity = 1.06-0.414 host activity. Host activity did not significantly influence vector activity (F1, 10 = 1.07; p>0.326).
Figure 6 shows the vector and host activity profiles at Nguruman during the dry season. Host activity showed two peaks, a morning peak at 0800 h and a prolonged mid day and afternoon high activity beginning as early as 1000 h and rising to a peak at 1400 h. The two prominent activity peaks of G. pallidipes coincide with the high activity period of the hosts. The highest interaction period between the vectors and the hosts occurs between 1000 and 1200 h in the morning and between 1600 and 1700 h in the evening. Regression analysis with vector as response and host as predictor variable defined the relationship as: Vector activity = 1.18+0.106 host activity. Host activity did not significantly influence vector activity (F 1, 10 = 0.27; p>0.616).
Wet Season Activity
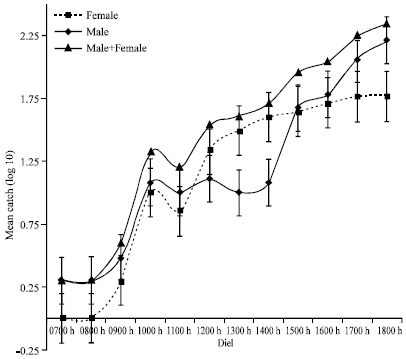
The activity of G. pallidipes at Busia during the wet season showed that the flies become active as from 0900 to 1700 h (Fig. 7). Males begun their activity earlier than females and became significantly hyperactive by about 1600 h. The activity of males declined sharply to zero by 1800 h. Females on the other hand exhibited delayed onset of activity in the morning h and a prolonged evening activity going beyond 1800 h. When both sexes were combined, the activity of G. pallidipes at Busia during the wet season showed a generalized unimodal pattern with a suppressed activity at about 1300 h. The morning activity of the flies intensified at 0900 h rising to attain the peak at 1200 h. The activity remained at peak until 1400 h, save for a slight suppression at 1300 h. The fly activity then decreased and maintained at a low rhythm to 1800 h. Contrarily the activity of both male and female G. pallidipes at Nguruman during the wet season showed increased catches with time throughout the day (Fig. 8). There was no major variation in activity between the sexes. A minor morning peak occurred at about 1000 h for both sexes after which males showed 2 h depressed activity before regaining high evening activity parallel to that of females. The evening peaks, which were the highest, occurred at 1800 h for both sexes. The activity profile for combined male and female G. pallidipes at Nguruman did not show any prominent peaks in the morning and afternoon during the wet season. The activity increased gradually with minor fluctuations throughout the day peaking at 1800 h. Flies started being active as early as 0700 h but vigorous activity was observed as from 1000 h. The highest peak was recorded at 1800 h. This profile is remarkably different from the dry season profile that showed prominent modes in morning and evening activity.
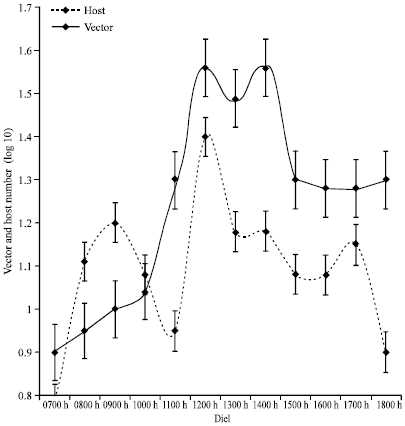
Figure 9 shows that the wet season activity of the vector and the hosts at Busia seem to coincide after 1000 h. The highest coincidence in activity occurs during the morning peak at 1200 h. Regression analysis with vector as response and host as predictor variable defined the relationship as: Vector activity = 0.441+0.739 host activity. Host activity did not significantly influence vector activity (F1, 10 = 3.6; p>0.087). However, the activity of the vector among the Nguruman sub-population followed that of the host at a one-hour lag between 0900 and 1200 h (Fig. 10). The activity of the vector steadily increased after midday, while that of the host oscillated at around the maximum level. Regression analysis with vector as response and host as predictor variable defined the relationship as: vector activity = 0.044+1.36 host activity. There was a significant influence of host activity on vector activity (F 10, 11 = 7.27; p<0.022).
Effect of Weather on the Activity of Vectors and Hosts at Busia and Nguruman
Effects of Weather on Vector and Host Activities at Busia
During the dry season, significant positive correlation between vector activity and temperature (r = 0.743, p<0.001) and between vector activity and lux (r = 0.479, p<0.018) was observed (Table 1). However, temperature had a stronger positive correlation to vector activity than lux. There was a significant negative correlation between vector activity and relative humidity (r = -0.428, p<0.037). Host activity correlated insignificantly to all weather variables. Lux and relative humidity correlated positively with host activity, while temperature correlated negatively to host activity. Vector activity had a weak and insignificant negative correlation with host activity (r = -0.102, p<0.635).
| Table 1: | Correlation analysis of temperature, relative humidity and lux on the activity of vector and host during the dry and wet seasons at Busia |
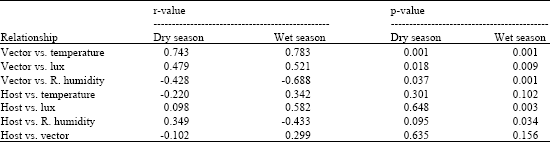 | |
| Table 2: | Regression analysis of temperature, relative humidity and lux on the activity of Glossina pallidipes during the dry and wet seasons at Busia |
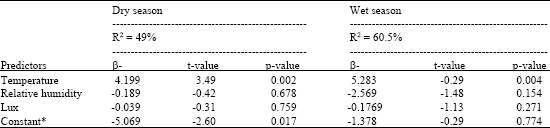 | |
| * Effect of factors not explained by the model | |
Weather factors significantly (F3, 20 = 8.37, p<0.001) influenced vector activity. Step-wise regression analysis resulted in regression equation log vector activity = -5.07+4.20 log temperature -0.040 log lux -0.189 log relative humidity, defining the influence of weather elements on vector activity. Weather factors explained 8.37% of total variation in vector activity and the predictive power of the model was about 50% (R2 = 49%) (Table 2). The effect of temperature on vector activity was significant (t = 3.49, p<0.002) and temperature was the most important predictor accounting for 80.45% of weather related vector activity. The influences of lux and relative humidity on vector activity were however insignificant. During the wet season, there was a significant positive correlation between vector activity and temperature (r = 0.783, p<0.001) and, between vector activity and lux (r = 0.521, p<0.009) (Table 1). Vector activity correlated negatively to relative humidity and the relationship was significant (r = -0.688, p<0.001). There was a weak positive correlation between host activity and temperature (r = 0.342, p>0.102), while lux showed a strong significant positive correlation to host activity (r = 0.582, p<0.003). The correlation between host activity and relative humidity was negative but significant (r = -0.433, p<0.034), while that between host and vector activity was weak and insignificant (r = 0.299, p>0.156). Step-wise regression analysis showed that temperature, lux and relative humidity significantly (F3, 20 = 12.76, p<0.001) influenced vector activity. The relationship between the weather factors and vector activity was captured in regression model Log vector activity = -1.38+5.28 log temperature -2.57 log relative humidity -0.177 log lux. The model fit was poor with weather factors explaining about 13% of total variation in vector activity. The predictive power of the model was however high (R2 = 60.5%), (Table 2). The influence of temperature on vector activity was significant (t = 3.27, p<0.004) and temperature was the most important predictor accounting for 65.82% of weather related vector activity. The influences of lux and relative humidity on vector activity were however insignificant.
Effects of Weather on Vector and Host Activity at Nguruman
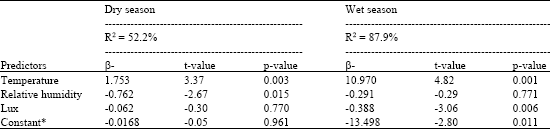
During the dry season, vector activity showed strong significant positive correlation to temperature (r = 0.66, p<0.001) and lux (r = 0.573, p<0.003). However, the correlation between vector activity and relative humidity was insignificant (r = 0.210, p>0.325) (Table 3). Host activity showed strong significant positive correlation with temperature (r = 0.527, p<0.008) and lux (r = 0.657, p<0.001). The correlation between relative humidity and host activity was however, negative and insignificant (r = 0.215, p>0.313). The correlation between host and vector activity was weak and insignificant (r = 0.156, p>0.466). Step-wise regression analysis showed that temperature, lux and relative humidity significantly (F3, 20 = 9.39, p<0.001) influenced vector activity. Regression model Log vector activity = -0.017+1.75 log temperature - 0.762 log relative humidity-0.062 log lux captured the influence of weather factors on vector activity. Weather factors explaining about 9% of total variation in weather related vector activity with the predictive power of the model being about 50% (R2 = 52.2%). Vector activity was significantly influenced by temperature (t = 3.37, p<0.003) and relative humidity (t = 2.67, p<0.015) and temperature was the most important predictor accounting for 68.5% of weather related vector activity (Table 4).
| Table 3: | Correlation analysis of temperature, relative humidity and light intensity on vector and host activity at Nguruman during the dry and wet seasons |
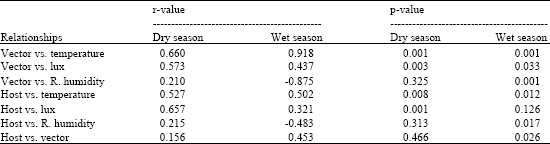 | |
| Table 4: | Regression analysis of temperature, relative humidity and lux on the activity of Glossina pallidipes during the dry and wet seasons at Nguruman |
 | |
| * Effect of factors not explained by the model | |
The influence of lux on vector activity was however insignificant.
During the wet season, vector activity showed strong significant positive correlation to temperature (r = 0.918, p<0.001) and a weak significant positive correlation to lux (r = 0.437, p<0.033). Vector activity however showed a strong significant negative correlation to relative humidity (-0.875, p<0.001). Host activity correlated significantly to temperature (r = 0.502, p<0.012), relative humidity (r = -0.483, p<0.017) and vector activity (r = 0.453, p<0.026) (Table 3). However, the correlation between host activity and lux was insignificant (r = 0.321, p>0.126). Step-wise regression analysis showed that temperature, lux and relative humidity significantly (F3, 20 = 56.78, p<0.001) influenced vector activity (Table 4). Regression equation Log vector activity = -13.5+11.0 log temperature -0.292 log relative humidity -0.388 log lux defined the model and showed good fit with weather factors explaining about 57% of total variation in vector activity. The predicative power of the model was about 90% (R2 = 87.9%). Temperature and lux showed significant effects, (t = 4.82, p<0.001) and (t = 3.06, p<0.006) respectively, on vector activity. However, temperature was the most important predictor among the weather factors accounting for 68.5%. The influence of relative humidity on vector activity was insignificant.
DISCUSSION
Both Busia and Nguruman subpopulations of G. pallidipes exhibited bimodal dry season activity for both sexes. However, a striking difference was observed in the onset of morning activity with the Nguruman sub-population having a much earlier onset than the Busia sub-population. The morning peaks were however more or less similar occurring between 1100 and 1200 h. Another major contrast in dry season activity of the two subpopulations was noted in the afternoon peaks. Whereas the Busia sub-population activity went to zero by sunset, that of Nguruman sub-population showed increased activity at dusk, an indication of twilight (crepuscular) behaviour. Crepuscular activity has been reported among the G. longipennis population at Galana ranch (Makumi et al., 1998). Vanderplank (1941) also observed the dusk activity behaviour in G. pallidipes in Tanzania. Wet season vector activity at Busia was unimodal, peaking earlier than that of dry season morning activity and showing a prolonged evening tail. The Nguruman activity profile during the wet season was unlike that in the dry season as it was unimodal and showed steady increase with time throughout the day. It however, compared well with that observed at Nguruman in terms of unimodal peak, only that the peaks showed up at completely different times of the day. The observed variations in activity patterns of subpopulations compare well to earlier observations on G. pallidipes. For example, Harley (1965) working in Busoga district of Uganda reported that there were no large seasonal changes in the activity pattern of G. pallidipes. Male flies showed a gradual increase in activity from early morning until evening, with a peak in the second hour before sunset, followed by a rapid fall. Peak female activity more closely coincided with maximum temperature than did male activity. Owaga et al. (1993) observed that the activity of G. pallidipes at the Kenyan coast was throughout the day with significant peaks of activity between 0900 and 1000 h and between 1400 and 1700 h.
In both vector subpopulations, diel activity correlated positively and significantly to temperature during dry and wet season. However, lux correlated positively and significantly to vector activity in both vector subpopulations during both seasons except the wet season at Nguruman. Step-wise regression analysis selected temperature as the key weather related factor accounting for 80.45 and 68.5% of vector related activities at Busia and Nguruman, respectively. Vanderplank (1941) similarly found temperature as the key regulator of G. pallidipes activity in Tanzania. Mohamed Ahmed and Odulaja (1997) also found temperature and light intensity as the best predictors of G. fuscipes catches around lake Victoria. Hargrove and Brady (1992) noted that high temperatures, (above 33°C) cause the normal activity pattern of tsetse to break down resulting in no late afternoon or evening peak of activity, this being replaced by a peak at dawn. This was precisely demonstrated among the Nguruman sub-population during the dry season when the mid day temperature soared to 38°C. However, low temperatures (below 24°C) results in the replacement of the dawn peak by a peak at dusk as is clearly demonstrated by the wet season activity of the Busia sub-population flies. It should however be noted that weather related factors accounted for only about 10% of vector activity in the two subpopulations, leaving a striking variation of about 90% in vector activity unaccounted for in each sub-population. The variation is likely to be espoused in host related activity and the innate genetic factors unique to the subpopulations.
Host movement correlated negatively and significantly to vector activity during the dry season but positively, though insignificantly, during the wet season among the Busia sub population. However, this was not the case at Nguruman where in both dry and wet seasons, host movement positively, though insignificantly, influenced vector activity. It should be noted that the diversity and abundance of host at Nguruman was much higher than that at Busia. Vectors therefore restrict their activity to high host activity periods since host searching is cheaper. Due to limited host availability and diversity at Busia, flies would spread their activity beyond that of hosts to maximize on chances of interaction. This explains less synchrony between host and vector activity patterns among the Busia as opposed to the Nguruman sub-population. The observed influence of host movement on vector activity explains part of the variations in vector activity that was not accounted for by the weather factors. It would be important to mention that in addition to environmentally stimulated activity, tsetse flies exhibit spontaneous activity that appear to result from an underlying circadian rhythm (Brady, 1970). Contact is a key epidemiological parameter in vector-transmitted diseases such as trypanosomiasis. The relationship between host and vector activity pattern highlights the nature and extent of contact between them. Synchronized activity pattern of vector and host increases the risk of transmission due to prolonged period of interaction.
ACKNOWLEDGMENTS
We are grateful to WHO/TDR, Swiss Tropical Institute (STI) and KARI-TRC for funding this work. We also thank Godfrey Emase, Joseph Etyang and Charles Nambiro for their participation in data collection during the field activities.
REFERENCES
- Brightwell, R., R.D. Dransfield, P. Stevenson and B. Williams, 1997. Changes over twelve years in populations of Glossina pallidipes and Glossina longipennis (Diptera: Glossinidae) subject to varying trapping pressure at Nguruman, South-West Kenya. Bull. Entomol. Res., 87: 349-370.
Direct Link - Makumi, J.N., C. Green and M. Baylis, 1998. Activity pattern of Glossina longipennis: A field study using different sampling methods. Med. Vet. Entomol., 12: 399-406.
PubMedDirect Link - Mohammed-Ahmed, M.M. and A. Odulaja, 1997. Diel activity patterns and host preferences of Glossina fuscipes fuscipes (Diptera: Glossinidae) along the shores of Lake Victoria, Kenya. Bull. Entomol. Res., 87: 179-186.
Direct Link