
Z. I.A. AL-Fifi
Department of Biological Sciences, Faculty of Science, King Abdulaziz University, P.O. Box 80094, Jeddah 21589, Saudi Arabia
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 4 | Page No.: 312-318







ABSTRACT
Dose-response studies of azadirachtin, dissolved in 90% aqueous ethanol and injected into freshly molted crowded fifth (last) instar nymphs of Schistocerca gregaria revealed three kinds of responses: (I) Delay of the next molt and death in the delayed molt, or after apolysis, without being able to escape from the old exuviae; (II) Production of over-aged nymphs surviving up to 83 days without molt or apolysis; (III) Death within 20 days after treatment, without molt, presumably due to a direct toxic effect of the azadirachtin. Solvent injected controls molted to adults within 8-10 days. With increasing doses, response I decreased and response III increased. A dose of 30.4 μg g-1 (μg azadirachtin per g fresh weight) resulted in response III in all nymphs. A dose of 15.2 μg g-1 induced the highest proportion of over-aged nymphs, but even with this dose only 13.1% over-aged nymphs were obtained. With doses lower or higher than 15.2 μg g-1, the proportion of over-aged nymphs was even lower. Over- aged male nymphs exhibited sexual behavior, courting and mounting over-aged female nymphs and adult females, without being able to copulate.
PDF Abstract XML References
How to cite this article
Z. I.A. AL-Fifi, 2006. Molting Inhibitory and Lethal Effects of Azadirachtin on the Desert Locust, Schistocerca gregaria (Forskal). Journal of Entomology, 3: 312-318.
DOI: 10.3923/je.2006.312.318
URL: https://scialert.net/abstract/?doi=je.2006.312.318
DOI: 10.3923/je.2006.312.318
URL: https://scialert.net/abstract/?doi=je.2006.312.318
INTRODUCTION
Azadirachtin, a limonoid found in the Neem tree (Azadirachta indica A. Juss), has antifeedant, growth disrupting (also termed molt-inhibitory or anti-molting) and lethal effects on insects (Jacobson, 1986; Aranson et al., 1989; Schmutterer, 1990, 1995; Ascher, 1993; Mordue (Luntz) and Blackwell, 1993; Xuan et al., 2004).
We demonstrated that a finely balanced dose of azadirachtin, administered by injection, inhibits the molt and induces long-surviving over-aged nymphs in the African migratory locust, Locusta migratoria migratorioides (Shalom and Pener, 1984, 1987). Other authors also reported on the molt-inhibitory effect of azadirachtin in L. migratoria (Sieber and Rembold, 1983; Mordue (Luntz) et al., 1985, 1986). Such over-aged nymphs of this species were utilized to study the interdependence between morphological and physiological metamorphosis; in other words, whether and how inhibition of morphological metamorphosis affects adult competence, that is physiological and behavioral patterns characteristic to adults (Shalom and Pener, 1984, 1987; Pener and Shalom, 1987; Shalom et al., 1988, 1993; Pener et al., 1989; Van der Horst et al., 1989; Wilps et al., 1992).
Although we recorded (Pener and Shalom, 1987) that injection of azadirachtin is capable of producing over-aged nymphs also in the desert locust, Schistocerca gregaria, no dose-response relations and their evaluation were published. The present article reports the dose-response data for producing over-aged nymphs in S. gregaria and distinguishes between the anti-molting and direct lethal effects of injected azadirachtin in this species of locusts.
MATERIALS AND METHODS
Insects
Crowded (gregarious) nymphs of Schistocerca gregaria from Ministry of Agriculture, Jeddah branch, Saudi Arabia were used. Experimental locusts were kept in 12-liter metal cages, 12-24 locusts per cage, under a 12:12 h light:dark regime, 38 (±1)°C during the photophase and 27 (±1) °C during the scotophase. The locusts were fed on Kikuyu grass (Pennisetum clandestinum Hochst), cabbage, both freshly supplied daily and flaked oats.
Administration of Azadirachtin and Experimental Design
Azadirachtin was dissolved in 90% aqueous ethanol in concentrations selected to comprise the desired dose, μg azadirachtin per g average fresh weight, in 2 μL of solvent. Such doses were injected, through the dorsolateral area of the intersegmental membrane between the third and fourth abdominal tergite, into fifth(= last)-instar crowded nymphs of S. gregaria, 1-24 h after the molt to this instar. Untreated and solvent injected controls were employed. The locusts were checked daily and molt to adult stage or mortality were noted. We distinguished between death in molt and death without molt. The former means that the insect died after complete apolysis; usually the old cuticle ruptured, but the locust were unable to escape from the exuviae and died in the course of an unsuccessful molt. In extreme cases of death in molt, the old cuticle only slightly ruptured, or did not rupture at all, but it was easy to peel the apolysed old cuticle. No apolysis took place in death without molt. If neither death, nor molt to the adult stage occurred and the locusts survived over 40 days, they were considered as over-aged nymphs. Observations on male sexual behavior of such over-aged nymphs were carried out and the weekly average percent of time spent on sexual behavior by the males was calculated (Pener and Lazarovici, 1979).
RESULTS
Figure 1 shows the percentage distribution of the different responses to injection of graded doses of azadirachtin. Mortality was higher in the untreated than in the solvent injected controls; therefore, these two groups of controls were pooled. Out of 48 control insects, 44 (91.7%, Fig. 1) molted to normal adults after spending 8-10 days in the fifth (= last) nymphal instar. The remaining four controls (8.3%, Fig. 1) died, not in molt, before reaching day 8. The cause of this mortality is unknown.
Following injection of the lowest dose, 1.9 μg g-1 (= 1.6 μg per nymph) of azadirachtin, 16.7% of the recipients (Fig. 1) molted to normal adults, or to adults with slightly curled wings, at the age of 10-12 days in the fifth nymphal instar. This is a slight delay in comparison to the controls. About 62% of the recipients of this dose died in the molt to adults; this is connoted as response type I. About 21% of the recipients died not in molt, connoted response type III (Fig. 1). With increasing doses, response I (death in molt) became less frequent, whereas the frequency of response III (death without molt) increased (Fig. 1). With doses as high as 30.4 or 121.6 μg g-1 of azadirachtin, solely response III was observed. Over-aged nymphs (response II, survival over 40 days in the fifth instar without molt) were obtained only with two doses, 7.6 and 15.2 μg g-1 of azadirachtin, yielding 7.4 and 13.1% over-aged nymphs, respectively (Fig. 1). Over-aged nymphs survived up to 83 days in the fifth instar.
With doses increasing from 1.9 to 15.2 μg g-1 of azadirachtin the timing of response I was increasingly belated, i.e., the locusts died in molt after spending more days in the fifth instar (Table 1). On the other hand, with increasing doses, the timing of response III shortened, death not in molt occurring earlier with the higher doses (Table 2).
| Table 1: | Dose-dependent timing of response I (death in molt) after injection of azadirachtin to 1 to 24 h old 5th instar nymphs of S. gregari |
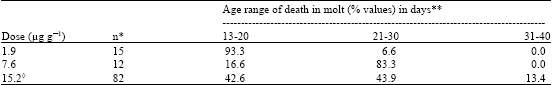 | |
| *n = number of fifth-instar nymphs which died in molt, **Percentage of fifth-instar nymphs which died in molt within the given age range, as based on n = 100% in the same line, ◊ No response I was observed with doses higher than 15.2 μg g-1 | |
| Table 2: | Dose-dependent timing of response III (death not in molt) after injection of azadirachtin to 1 to 24 h old 5th instar nymphs of S. gregaria |
 | |
| + Data are not shown for the dose of 1.9 μg g-1 because only five nymphs died not in molt and this small value of n does not justify presentation in percent values, * n = number of fifth-instar nymphs which died not in molt, ** % of fifth-instar nymphs which died not in molt within the given age-range, ◊ All died within 2 days after injection of azadirachtin | |
Dose-dependent timing of response I (death in molt) after injection of azadirachtin to 1 to 24 h old fifth-instar nymphs of S. gregari.
Dose-dependent timing of response III (death not in molt) after injection of azadirachtin to 1 to 24 h old fifth-instar nymphs of S. gregaria.
Observations on male sexual behavior of the over-aged nymphs were carried out for a three week period, 40-60 days (both included) after the molt to the fifth instar.
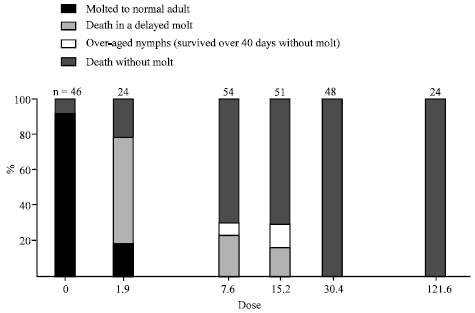 | |
| Fig. 1: | Effects of graded doses of azadirachtin injected to 1 to 24 h old 5th instar-nymphs of S. gregaria. Abiscisse (in logarithmic scale): dose injected (g azadirachtin per g average fresh weight). Ordinate: percent value (indication distributions of the different responses). The number of nymphs (n) injected with a given dose is shown on the top of the respective column; this number constitutes 100% for that column. Dose O marks controls |
 | |
| Fig. 2: | Fifth instar over-aged male-nymph of S. gregaria exhibiting sexual behavior toward over-aged female nymph |
In two separate groups of over-aged male nymphs, the weekly average percentage of time spent on sexual behavior varied between 6 to 10.5 % (n = 8 males in each group for each week). Over-aged male nymphs exhibited sexual behavior towards both, over-aged female nymphs (Fig. 2) and untreated adult females; they courted and mounted the females and bent their abdomen to reach the females’ genitalia, but were unable to copulate. Over-aged male nymphs, like normal adult males, also mounted fellow males, especially in absence of females. Observations carried out before day 40 showed that male sexual behavior initiated at the age of 33 and 36 days in the two groups, respectively, after the molt to the 5th instar.
DISCUSSION
The results confirm the molt-inhibitory effect of azadirachtin in Schistocerca gregaria. Following treatment of young fifth-instar nymphs, we found that with increasing doses the response changed from slightly delayed but still successful molt (considered as a border case, but still normal development) to death in a more delayed unsuccessful molt (response I, death in molt or after apolysis without shedding the exuviae), then to long-surviving over-aged nymphs (response II) and finally to death not in molt (response III). Some of these responses markedly overlapped after administration of the lower doses, but in the cases of higher doses (30.4 and 121.6 μg g-1) only response III was observed.
Although both response I and III are lethal effects of azadirachtin, these two kinds of mortality are basically different. Response I should be considered as a physiological death, corresponding with the molt-inhibitory effect of the azadirachtin. The reason why delay in the molt coincides with inability of shedding the exuviae is unknown. In contrast, response III seems to be caused by a toxic effect of azadirachtin, not related to its molt-inhibitory effect because with higher doses it occurs before the molt becomes actual. With the highest dose administered, 121.6 μg g-1 (Table 2), all nymphs showed response within two days after treatment, that is they died on the second and third day after molt to the fifth instar, about a week before the next molt in normal (control) locusts. In addition to its antifeedant and molt-inhibitory effects, azadirachtin is known to affect many organs and physiological systems in insects (Mordue (Luntz) and Blackwell, 1993) and some of these may result in lethal-toxic consequences.
Over-aged nymphs (response II) were obtained only within a narrow range of doses and the proportion of the over- aged nymphs was low even with such doses. Response II always co-occurred with both, response I and III, but response III was dominating in these cases (Fig. 1).
The presently reported dose-response relations of S. gregaria to azadirachtin are roughly similar to those found in previous studies for Locusta migratoria (Shalom and Pener, 1987; Pener, 1987; Wilps et al., 1992). With increasing doses, both species show a shift from response I, through response II, to response III. However, S. gregaria seems to be somewhat less susceptible to azadirachtin. The dose which induces maximum proportion of over-aged nymphs is over five times higher in S. gregaria (15.2 μg g-1) than in L. migratoria (2.9 μg g-1 = 1.6 μg per fifth-instar nymph). Also, in S. gregaria, the lowest dose to induce response III in all recipients is 30.4 μg g-1 (Fig. 1), whereas in L. migratoria it is only 11.4 μg g-1 (=6.4 μg per nymph). Another minor difference between the two species is that with the optimal dose for inducing over-aged nymphs, a slightly lower proportion of such nymphs are obtained in S. gregaria (13.1%) than in L. migratoria (19.4 %) (Shalom and Pener, 1987).
Azadirachtin-induced over-aged male nymphs of S. gregaria exhibited male sexual behavior. However, the intensity of this behavior was relatively low. The values of the weekly average percentage of time spent on sexual behavior by the over-aged male nymphs (6-10.5%,) were only about one-fifth to one-tenth of the values obtained for normal adult males (50-60%), kept under similar condition and submitted to the same method of observations (Pener and Lazarovici, 1979).
Langewald and Schmutterer (1992, 1995) reported that treatment of crowded S. gregaria nymphs with neem seed oil, or with neem seed oil enriched by azadirachtin, induced a phase shift toward solitary direction. These authors found (among other findings) that the proportion of nymphs showing solitary coloration is higher in the treated than in the untreated locusts. In our experiments on the effect of azadirachtin on crowded S. gregaria nymphs, we observed no color change toward the solitary phase. However, our experimental design was very different from that of Langewald and Schmutterer (1992, 1995). The dose of azadirachtin-enriched neem oil, applied by Langewald and Schmutterer (1995) to second and third-instar nymphs of S. gregaria, was certainly low, because it allowed molt to the fourth and fifth instar. The effect of azadirachtin, or of neem extracts, as well as of seed-kernel extracts from Melia volkensii (Nasseh et al., 1993), on locust phase polymorphism is somewhat problematic (Pener and Yerushalmi, 1998), but clearly deserves further investigations. Azadirachtin seems to inhibit release of neurosecretion from the corpora cardiaca Subrahmanyam et al., 1989) and some other limonoids may exert a similar effect. Therefore, it cannot be ruled out that the relatively low doses administered by Langewald and Schmutterer (1995) interfered to some extent with the release of the newly discovered dark-color- inducing neurohormone of locusts (Tawfik et al., 1999), eventually resulting in diminution/disappearance of the gregarious dark patterns in the nymphs after one or more molts.
REFERENCES
- Ascher, K.R.S., 1993. Nonconventional insecticidal effects of pesticides available from the Neem tree, Azadirachta indica. Arch. Insect Biochem. Physiol., 22: 433-449.
CrossRefDirect Link - Jacobson, M., 1986. The Neem tree: Natural resistance par excellence. A.C.S. Symposium Series, 296: 220-232.
Direct Link - Mordue, A.J. and A. Blackwell, 1993. Azadirachtin: An update. J. Insect Physiol., 39: 903-924.
Direct Link - Pener, M.P., T.M. van den Broek, W.J.A. van Marrewijk, J.M. van Doorn, D.J. van der Horst and A.M.T. Beenakkers, 1989. Development of imaginal competence to adipokinetic hormone in Locusta: Lipid and carbohydrate levels and glycogen phosphorylase activity in azadirachtin-induced over-aged nymphs. Comparat. Biochem. Physiol., 94B: 293-298.
Direct Link - Schmutterer, H., 1990. Properties and potential of natural pesticides from the neem tree, Azadirachta indica. Annu. Rev. Entomol., 35: 271-297.
CrossRefDirect Link - Subrahmanyam, B., T. Muller and H. Rembold, 1989. Inhibition of turnover of neurosecretion by azadirachtin in Locusta migratoria. J. Insect Physiol., 35: 493-500.
CrossRefDirect Link - Van der Horst, D.J., J.M. van Doorn, M.P. Pener, A.T.M. van den Broek, W.J.A. van Marrewijk and A.M.T. Beenakkers, 1989. Development of imaginal competence to adipokinetic hormone in Locusta: Lipophorin conversions in precocene- induced adultiforms and in azadirachtin-induced over-aged nymphs. Comparat. Biochem. Physiol., 92B: 133-136.
Direct Link - Xuan, T.D., T. Eiji, T. Hiroyuki, M. Mitsuhiro, T.D. Khanh and I.M. Chung, 2004. Evaluation on phytotoxicity of neem (Azadirachta indica. A. Juss) to crops and weeds. Crop Prot., 23: 335-345.
CrossRef