
Hazem S. Hasan
Instructor in Plant Protection, Department of Agricultural Sciences, Al-Shobak University Collage, Al-Balqa Applied University, I 9I I 7-Salt-Jordan
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 4 | Page No.: 290-297







ABSTRACT
Random amplified polymorphic DNA polymerase chain reaction (RAPD-PCR) was used to survey the B biotype and other biotypes of Bemisia tabaci in Jordan. Whiteflies were collected from cultivated plants, wild plants and weeds from (9) different localities and on (12) distinct crops with total (123) whitefly insect. RAPD analyses using three selected 10-mer primers reliably identified B. tabaci biotypes. The total number of clear bands obtained from it was 29 DNA bands. Cluster analysis demonstrated that, in general, biotype B individuals which are scattered independently in the localities according to the host plant and coexists with cultivated plants. While an intermediate whitefly populations BA having biotype A a distinguish band only with OPR-04 but according to Jaccard similarity they have higher genetic distance with biotype B and this samples where restricted to wild plants and weeds. And finally the biotype A was restricted to the isolated area comported to other biotype. The percentage of Whitefly biotype A, B and BA in the sample was approximately 12.5, 75 and 12.5% Jordan Valley and Upland of tested samples.
PDF Abstract XML References
How to cite this article
Hazem S. Hasan, 2006. Survey of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) Biotypes in Jordan using RAPD marker. Journal of Entomology, 3: 290-297.
DOI: 10.3923/je.2006.290.297
URL: https://scialert.net/abstract/?doi=je.2006.290.297
DOI: 10.3923/je.2006.290.297
URL: https://scialert.net/abstract/?doi=je.2006.290.297
INTRODUCTION
Whiteflies of the Bemisia tabaci (Gennadius) group are a major and increasing problem for agriculture throughout the world (Blackman and Cahill, 1998; Perring, 2000). The problem involves the recognition of a complex of sibling species, host races and/or biotypes which vary in their life-history parameters (Drost et al., 1998), virulence (Brown et al., 1995) and in their response to the same chemical (Sharaf, 1986). This variability stimulated intensive multidisciplinary research on this insect in recent years (Gerling and Mayer, 1996).
Different populations of B. tabaci differ in their phenology, ability to utilize particular host-plants and virus transmission characteristics (Burban et al., 1992; Bedford et al., 1994; Markham et al., 1996), but they are indistinguishable based on morphological characteristics (Mound and Halsey, 1978; Rosell et al., 1997). A more recent realization is that there is a high degree of genetic heterogeneity between different B. tabaci populations, either from different geographical areas or different host-plant species at the same site (Burban et al., 1992; Bedford et al., 1994; Brown et al., 1995).
Analysis of isozyme variation in esterases in world-wide B. tabaci populations revealed the existence of considerable genetic variation. Based on esterase profiles, new biotypes were proposed and currently 20 of them, named A-S are recognized (Brown et al., 1995; Rosell et al., 1997; Banks et al., 1999). In 2002, the poinsettia strain, silverleaf whitefly or B biotype of Bemisia tabaci was detected in Jordan (Sharaf and Hasan, 2003).
RADP-PCR is a relatively simple, inexpensive and rapid technique, revealing polymorphisms which are useful as genetic and taxonomic markers (Lima et al., 2000; Zhang et al., 2005). RAPD has been applied to study of insects and to differentiate whiteflies, including the identification of the A and B biotypes of B. tabaci (Loxdale and Lushai, 1998 and Gawel and Bartlett, 1993).
The present study is an attempt to demonstrate the use of RAPD-PCR to denote the presence of B. tabaci biotypes in Jordan; Determine the different host plants for whitefly in Jordan during the studded period. Identify the collected samples according to the random amplified polymorphic DNA PCR (RAPD-PCR) fingerprinting technique. The objectives of the present study were to use RAPD markers (1) to estimate the genetic diversity of B. tabaci populations, (2) to discriminate between B. tabaci biotypes.
MATERIALS AND METHODS
Insects Collection
Adult whiteflies were collected from host plants and fields using a hand-held aspirator or forth larval stage (Pupae); which reared to reach adult stage during the period from October 2004 to May 2005 from the Jordan Valley and some area from Upland. A total of 17 samples (Table 1), each of approximately 123 insects, were collected from 5 locations in Jordan and on 22 distinct crops. All insects used in this study were from infestations identified as B. tabaci based on morphology of the pupal stage. Reference colonies of A biotype adults originating from JU culture. DNA was extracted from 5 individual insects of each sample, according to a standard protocol with several modifications.
DNA Extraction
DNA was extracted from individual insect of each sample were modified slightly from those described by Black et al. (1992) by using Wizard® Genomic DNA Purification Kit (Promega) as mentioned by Sharaf and Hasan (2003).
RAPD Assays and Electrophoresis
The 10 mer primers (Operon Technologies Inc., Alameda, CA, USA) used for RAPD-PCR are shown in Table 2. PCR was performed in a 30 μL reaction mix. Samples were amplified using a thermocycler with the following program: one cycle of 3 min at 94°C followed by 45 cycles of 1 min at 93°C, 1 min at 35°C and 2 min at 72°C, followed by a final cycle of 5 min at 72°C.
| Table 1: | Host plants, sites where whitefly populations were collected, population codes used in the analyses |
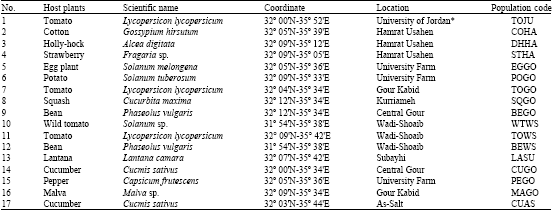 | |
| * Known species as Biotype A Bemisia tabaci from University of Jordan (JU) | |
| Table 2: | Sequence of the primers OPA-4, OPA-11 and OPA-17 |
PCR products were separated by electrophoresis on 1.5% agarose gels. A 100 bp ladder (Amersham Pharmacia Biotech) was included on each gel as molecular weight marker. DNA bands were visualized under UV light after staining with a 0.5 μg mL-1 ethidium bromide solution. Gels were documented using the Gels Documentation System. A negative control without DNA was included in all reaction sets.
Data scoring and Analysis
PCR amplification products of the 123 samples were scored as presence or absence of bands. The data matrix was used to calculate Jaccard’s similarity coefficient, which does not consider the joint absence of a marker as an indication of similarity. A dendrogram was constructed using the Unweighted Pair-group Method Analysis (UPGMA). The frequencies of the RAPD fragments were estimated for each of the 15 Bemisia populations and for the two other whitefly species. The resulting matrix was used to calculate Manhattan distances between all pairs of populations or species. These analyses were performed using SPSS-pc software, version 11.0.
RESULTS
RAPD-PCR was used to identify whitefly B. tabaci biotypes from 9 sampling sites originating different location in Gour and upland and different host plants. Representative sample of B. tabaci biotype A. collected from Tomato cultured in University of Jordan rearing room (TOJU) were included in each reaction set as identification control. Primers screened produced clear bands on the RAPD amplifications and were used for analysis. These primers (OPA-04, OPA-10 and OPA-17) amplified a total of 29 markers. The total number of clear bands obtained from each primer is 9 (OPA-04) to 12 (OPA-10) and 8 (OPA-17) and with an average of 9.6 bands per primer. The size of amplification products ranged from 160 to 1000 bp. RAPD amplification patterns are shown in Fig. 1. All primers produced identical RAPD patterns for the selected bands in the primer screening step and in the final analysis and in the several replicated experiments that were done.
Distribution of B. tabaci biotype A was dominant at cotton Hamrat Usahen (COHA) and tomato Gour Kabid (TOGO), the genetic distance of these two sample with (TOJU) equal to 0.417-0.700, respectively, that mean 2 out of 16 (exclude University of Jordan). On the other hand the area from central Gour and North was dominant with B. tabaci biotype B demonstrated by the reaming 14 samples according to the Table 3 with genetic distance values referred to (TOJU) ranging from 0.056 to 0.067. Unless two of this 14 sample, wild tomato (WTWS) and Holly-hock (DHHA) have smaller genetic distance with (TOJU) equal to 0.200-0.111, respectively, on the other hand (DHHA) show the biotype A distinguished band in primer OPA-04 and OPA-017 and (WTWS) show this band in only OPA-04, while they show biotype B bands on the other primers, so we can consider them an intermediate population between biotype A and B with more relation to biotype B and can be represented as (BA).
| Table 3: | Proximity Matrix between B. tabaci populations. Population codes listed in Table 1 |
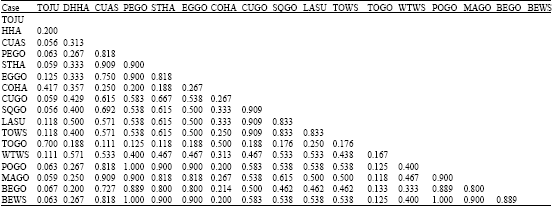 | |
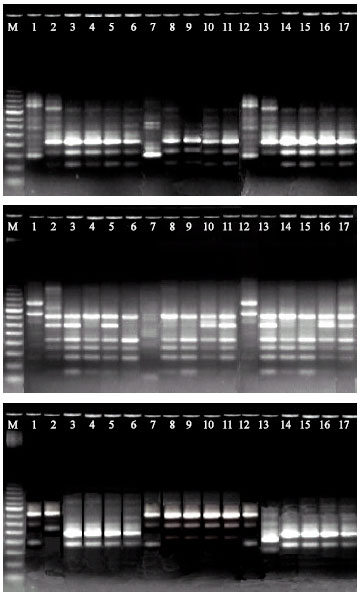 | |
| Fig. 1: | RAPD patterns generated by OPA-04 (a), OPA-10 (b) and POA-17 (c) Population codes listed in Table 1 |
The percentage of Whitefly biotype A, B and BA in the samples were approximately 12.5, 75 and 12.5%, respectively Jordan Valley and Upland.
An UPGMA dendrogram based on Jaccard’s similarity coefficient was constructed for the 123 analyzed samples. Two major groups were evident. The first group contains all populations of B. tabaci biotype A from different localities (Fig. 2).
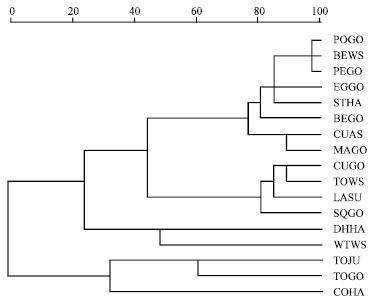 | |
| Fig. 2: | Dendrogram using average linkage (between groups) for 123 Whitefly samples, generated by UPGMA, Population code are listed in Table 1 |
This group is basically divided into tow subgroups. The first subgroup consists of individuals collected on cotton, from Hamrat Usahen (COHA). The second includes tomato from Gour kabid (TOGO). The reference culture (TOJU) tomato plants from university of Jordan. The second major group included populations of B. tabaci biotype A and B. It was divided to three subgroups, the first one include populations collected from wild plants from Wadi Shoaib (WTWS) and Hamrat Usahen (DHHA). While the second one was collected mainly from cultivated plants collected from As-Salt (CUAS), Wadi Shoaib (BEWS and TOWS), central Gour (BEGO). University Farm (PEGO, EGGO and POGO), Hamrat Usahen (STHA) and Ghour Kabid (MAGO) and in between this two subgroup is the third subgroup which demonstrated farm collected plants scattered in locations; Subayhi (LASU), Kurriameh (SQGO), Wadi Shoaib (TOWS) and Central Gour (CUGO).
DISCUSSION
A significantly change in the productivity of B. tabaci females has been documented during the 1990’s (Abu-Shribi, 1996; Hasan, 1999) as compared to that reported in 1980's (Batta, 1984; Sharaf and Batta, 1985). Perring (2001) suggested Bemisia tabaci was a species complex with many biotypes. Up to now, approximately 24 biotypes have been identified and characterized to different degrees. It was primarily divided into B biotype, Q biotype and non-B/Q biotype. The non-B/Q biotype includes more than 20 biotypes, such as A, K, D, E, G, H, L, M, N, etc. (Brown et al., 1995; De Barro et al., 1998; Perring, 2001). The objective of this study was to analysis the genetic variation of B. tabaci population in relation to their degree of genetic isolation and their pattern of distribution in the Jordan Valley. RAPD-PCR experiments show biotypes differed in the number and molecular weight of the produced bands (ranged from 160 to 1000 bp.) and (55.8% similarity) (Fig. 1 and Table 3). This agree with what found in 2002 (Sharaf and Hasan, 2003) in Jordan and over different places in the world (Moya et al., 2001; Zhang et al., 2005).
A surprising result produced by the cluster analysis (Fig. 2) was the grouping of B. tabaci biotypes A and B the second group. This support the suggestion that we may have also other biotypes rather than A and B, since we have a clear branching in the dendrogram with an approximately 75% biotype B in the sample. These results give support to the preliminary studies based on allozymes (Perring et al., 1992; Bellows et al., 1994), RAPD, crossing experiments and mating behavior studies (Perring et al., 1993) that had led the authors to conclude that B. tabaci biotypes A and B could be different species. Although data are insufficient to support the raising of any biotype to new species status, the fact remains that B. tabaci biotypes A and B are not a genetically similar. And also we have a genetic distance between B. tabaci biotype B.
Cluster analysis showed that in general biotype B individual are scattered independently in the locations where the samples were collected joining individuals that according to the host plants. This result suggests that a differentiation of population has already occurred, mainly according to the host plants, instead of the geographical region where populations are localized. No grouping of samples collected on different crops in the same location was obtained. For examples individuals collected on three different crops (Wild tomato, Tomato, Bean) from same location Wadi Shouib did not cluster according to the locality but to the crop (Dendrogram) but individuals collected from two very distant locations Gour kabid and salt (CUGO) and (CUAS) but on same crop cucumber were joined in the dendrogram. The cluster analysis of biotype B population showed the heterogeneous set of groups and subgroup, probably due to differences between several nutritional and behavioral factors; massive and crop-specific insecticide applications in different crops or by possible crossings between Whitefly biotypes individuals and indigenous biotype individuals. These results suggest that management methods established for a host crop in one region would probably function for the same crop in other regions.
The pure population of biotype A in Hamrat Usahen and the biotype B which found on cultivated plants and the biotype BA on wild plants and weed with relation to virus infection notice on host plants this may agree with what Al Musa (1986) found that in spite of absent of planted tomatoes plants around the year the infection of Tomato Yellow Leave Curl Virus (TYLCV) remains the most important factor limiting production of fall growing tomatoes in Jordan. Since this virus is transmitted by B. tabaci Biotype B (according to the result) that over summer on cultivated crops in side valleys or as Biotype BA (according to the result) on wild weeds or ornamental in Jordan valley providing a reservoir for the virus as well as vector. As conclusion we have in Jordan at least 3 biotype which are biotype A, B and BA.
Recent phylogenetic studies using more effective markers, such as mitochondrial DNA (Frohlich et al., 1999; Kirk et al., 2000) and ribosomal DNA markers (De Barro et al., 1998), showed that the renaming of the B biotype to B. argentifolii is premature. These studies suggest that the best way to view B. tabaci is as a complex containing geographically distinct populations that exhibit variation across a number of traits. More studies are necessary concerning molecular sequences (mitochondrial and nuclear genomes), the mechanism that regulate competition between the biotypes, host range phenotypes and mating compatibility among biotypes and sub-groups of B. tabaci to elucidate this controversy.
ACKNOWLEDGMENTS
The authors wish to express his gratitude to Dr. M. Sadder, for his invaluable comments and input on an earlier draft. Also I wish to thank Prof. Ahmad Katbeh and Instructor Leena Irshaid for technical assistance and for their help in writing the manuscript.
REFERENCES
- Bedford, I.D., R.W. Briddon, J.K. Brown, R.C. Rosell and P.J. Markham, 1994. Geminivirus transmission and biological characterisation of Bemisia tabaci (Gennadius) biotypes from different geographic regions. Ann. Applied Biol., 125: 311-325.
CrossRefDirect Link - Bellows Jr., T.S., T.M. Perring, R.J. Gill and D.H. Headrick, 1994. Description of a species of Bemisia (Homoptera: Aleyrodidae). Ann. Entomol. Soc. Am., 87: 195-206.
CrossRefDirect Link - Brown, J.K., D.R. Frohlich and R.C. Rosell, 1995. The sweetpotato or silverleaf whiteflies: Biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol., 40: 511-534.
CrossRefDirect Link - Burban, C., L.D.C. Fishpool, C. Fauquet, D. Fargette and J.C. Thouvenel, 1992. Host associated biotypes within West African populations of the whitefly Bemisia tabaci (Genn.) (Homoptera, Aleyrodidae). J. Applied Entomol., 113: 416-423.
Direct Link - De Barro, P.J., W. Liebregts and M. Carver, 1998. The distribution and identity of biotypes of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in member countries of the secretariat of the pacific community. Aust. J. Entomol., 37: 214-218.
CrossRefDirect Link - Frohlich, D.R., I. Torres-Jerez, I.D. Bedford, P.G. Markham and J.K. Brown, 1999. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol. Ecol., 8: 1683-1691.
CrossRefPubMedDirect Link - Gawel, N.J. and A.C. Bartlett, 1993. Characterization of differences between whiteflies using RAPD-PCR. Insect Mol. Biol., 2: 33-38.
PubMedDirect Link - Kirk, A.A., L.A. Lacey, J.K. Brown, M.A. Ciomperlik and J.A. Goolsby et al., 2000. Variation in the Bemisia tabaci s. 1. species complex (Hemiptera: Aleyrodidae) and its natural enemies leading to successful biological control of Bemisia biotype B in the USA. Bull. Entomol. Res., 90: 317-327.
Direct Link - Lima, L.H.C., D. Navia, P.W. Inglis and M.R.V. De Oliveira, 2000. Survey of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) biotypes in Brazil using RAPD markers. Genet. Mol. Biol., 23: 781-785.
Direct Link - Markham, P.G. I.D. Bedford, S. Liu, D.F. Frohlich, R. Rosell and J.K. Brown, 1996. The Transmission of Geminiviruses by Biotypes of Bemisia tabaci (Gennadius). In: Bemisia 1995: Taxonomy, Biology, Damage, Control and Management, Gerling, D. and R.T. Mayer (Eds.). Intercept Ltd. and Over, UK., pp: 69-75.
- Moya, A., P. Guirao, D. Cifuentis, F. Beitia and J.L. Cenis, 2001. Genetic diversity of Iberian populations of Bemisia tabaci (Hemiptera: Aleyrodidae) based on random amplified polymorphic DNA-polymerase chain reaction. Mol. Ecol., 10: 891-897.
CrossRefDirect Link - Perring, T.M., A.D. Cooper and D.J. Kazmer, 1992. Identification of the poinsettia strain of Bemisia tabaci (Homoptera: Aleyrodidae) on broccoli by electrophoresis. J. Econ. Entomol., 85: 1278-1284.
Direct Link - Perring, T.M., 2001. The Bemisia tabaci species complex. Crop Prot., 20: 725-737.
CrossRefDirect Link - Perring, T.M., A.D. Cooper, R.J. Rodriguez, C.A. Farrar and T.S. Bellows Jr., 1993. Identification of a whitefly species by genomic and behavioral studies. Science, 259: 74-77.
CrossRefPubMedDirect Link - Rosell, R.C., I.D. Bedford, D.R. Frohlich, R.J. Gill, J.K. Brown and P.G. Markham, 1997. Analysis of morphological variation in distinct populations of Bemisia tabaci (Homoptera: Aleyrodidae). System, 90: 575-589.
Direct Link - Sharaf, N., 1986. Chemical control of Bemisia tabaci. Agric. Ecosys. Environ., 17: 111-127.
CrossRefDirect Link - Zhang, L.P., Y.J. Zhang, W.J. Zhang, Q.J. Wu, Q.J. Xu and D. Chu, 2005. Analysis of genetic diversity among different geographical populations and determination of biotypes of Bemisia tabaci in China. J. Applied Entomol., 129: 121-128.
Direct Link