
A. Suganthi
Department of Agricultural Entomology, Tamil Nadu Agricultural University, Coirnbatore-641 003, Tamil Nadu, India
S. Chandrasekaran
Department of Agricultural Entomology, Tamil Nadu Agricultural University, Coirnbatore-641 003, Tamil Nadu, India
A. Regupathy
Department of Agricultural Entomology, Tamil Nadu Agricultural University, Coirnbatore-641 003, Tamil Nadu, India
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 3 | Page No.: 236-240







ABSTRACT
The bud worm, Hendecasis duplifascialis Hampson is one of the major pests of jasmine, Jasminum sambac. The pheromone release behaviour of the female was studied by making visual observations under a 12: 12D photoperiod. Most of the females (85.00%) called two days after emergence and majority of the females had multiple bouts of calling. The earliest initial calling was at 04.20 h. after the commencement of scotophase. The termination of calling was found to be as late as 10 min after the onset of photophase. Wind tunnel bioassay showed that two day old females were found to be more attractive to males and attraction was more pronounced in the presence of air current.
PDF Abstract XML References
How to cite this article
A. Suganthi, S. Chandrasekaran and A. Regupathy, 2006. Pheromone Release Behaviour of Jasmine Bud Worm, Hendecasis duplifascialis Hampson (Pyraustidae: Lepidoptera). Journal of Entomology, 3: 236-240.
DOI: 10.3923/je.2006.236.240
URL: https://scialert.net/abstract/?doi=je.2006.236.240
DOI: 10.3923/je.2006.236.240
URL: https://scialert.net/abstract/?doi=je.2006.236.240
INTRODUCTION
Jasmine (Jasminum sambac) known as the king of flowers, is one of the oldest fragrant flowers cultivated by man. It is a key Forex Earner. Essential oil extracted from jasmine flowers is used in perfume industry. The demand for flower extracts for perfume trade is growing. The major bottleneck in production of flower buds is the ravage by insect pests. Among the different insect pests recorded, bud borer Hendecasis duplifascialis Hampson (Lepidoptera: Pyraustidae) (David, 1958) is known to pose a serious threat to flower production. It causes damage to the tune of 40-50%, affecting the quality of the flowers and attributes for 30-70% yield loss (Gunasekaran, 1989). Amutha (1994) studied the population fluctuation of H. duplifascialis on five cultivars of jasmine and noted the prevalence of the insect throughout the year (Amutha, 1994). Currently bud worm control relies mainly on the use of insecticides which carries the undesired side effects of insecticide resistance, residues and reduction in beneficial natural enemies. Hence, there is a need to go for alternative methods of pest control.
Sex pheromone production in jasmine pests remains unexploited. A solid understanding of the pheromone biology, for the species in question, is essential for exploitation of pheromones. In view of the utility of sex pheromones for control of jasmine budworm, a study was made on the calling behaviour and temporal pattern of pheromone release of H. duplifascialis.
Materials and Methods
Mass Culturing of Test Insect
Field collected H. duplifascialis larvae were reared on jasmine buds using plastic tray kept under mosquito net. Fresh unopened buds were provided every day. Pupae were collected and sexed based on the difference in the terminal segment. Female pupa has genital operture located (dark slit like marking) on the eighth abdominal segment whereas, in male pupa, it is seen on the anterior region of ninth abdominal segment. Ten pairs of newly emerged male and female moths were introduced into rearing cages (0.6x0.3x0.45 m). Cotton pad soaked in 10% sugar solution was provided as adult food. The substrate contained two plastic tea stainer put in a juxta position (0.08 m diameter) containing jasmine buds as an odour source for oviposition and it was wrapped with white khada cloth. This was then hung from the top of the cage. The eggs laid on the cloth were used for further culturing of bud worm.
Calling Behaviour
For behavioural observation, immediately after emergence virgin females were individually caged in plastic containers (0.15x0.07 m) and were provided with sugar soaked cotton pad as adult food. Visual observations on characteristic posture resembling calling by female were recorded at 30 min intervals during the scotophase (dark phase), as the insect is reproductively active during this period. Ten each of freshly emerged, one, two and three days old females were put under constant observation to find out whether all females initiate calling. Experiment was repeated six times. Observations on calling, bout (one period of continuous calling) and bout length were recorded. Observations during the scotophase continued even if a female stopped calling. During photo phase, when no female called for a period of one hour, observations were discontinued.
Sex Pheromone Attraction Study
Sex pheromone attraction studies were conducted with an wind tunnel used by Sundararaju et al. (1994). with a slight modification. The wind tunnel was fabricated with transparent mylar sheet. The sheet was folded into a tubular shape (1 m length and 0.07 m diameter). Two transparent perforated plastic containers, one designated as bait and second as test chamber were attached one on either side of the open ends of the tunnel. Each chamber had a removable lid at the top and two circular holes (0.07 m in diameter) on opposite sides. The mouth of the tunnel at the bait chamber was covered with muslin cloth to confine the insects to the bait chamber. The holes at the free ends were covered with nylon net to allow wind movement. Cotton pad was soaked with sugar solution and placed over mouth of the container to serve as food source in both the chambers.
A power operated mini-table fan with electronic regulator was kept in front of the hole of bait chamber to carry air current from the bait chamber to the test chamber. The male and female were sexed based on pupal character. Sex pheromone attraction was assessed by keeping group of 15 individuals of one, two and three days old females separately in bait chamber and males in test chamber. The experiment was repeated with opposite sex in test and bait chamber and the number of insects reaching the bait chamber was counted during scotophase, with the help of a torch light covered with red gelatin paper. A set of blank experiment was run keeping only food source in bait chamber. Experiment was repeated six times.
RESULTS AND DISCUSSION
Visual observations revealed that the receptive H. duplifascialis females exhibited a typical calling posture. During calling, the terminal abdominal segments were slowly extended and retracted at regular intervals and wing fluttering was observed. The calling behavior was less observed in females that emerged within 24 h (0-day-old). As their age progressed, the percent calling females increased and the maximum percentage was recorded in two day-old females (85.00%) (Table 1).
| Table 1: | Calling behaviour of virgin adult females of H. duplifascialis |
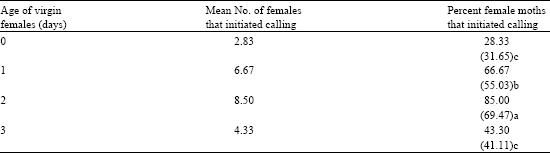 | |
| Figures in the parentheses are arc sine | |
| Table 2: | Pheromone response of H. duplifascialis (Adult moths) |
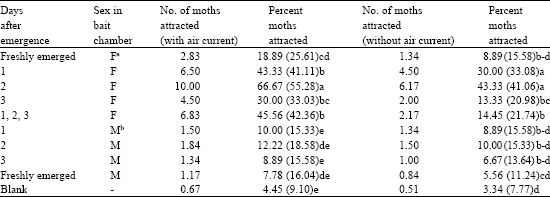 | |
| a- Female,b-Male, Figures in the parentheses are arc sine | |
In Earias vitella, Tamhankar et al. (1993) found that 83.3 and 96.9% of the females initiated calling on days 1 and 2 after emergence. This behaviour was more or less common to other moths like Palpita unionalis (Mazomenos et al., 2002) and E. insulana (Tamahankar, 1995).
Out of 20 two day old females, 11 had 2-5 bouts of calling, 9 had only single bout. Once the calling behaviour in female started, this posture continued for nearly 1.5 h. The bout lengths varied from 14-72 min for females calling in multiple bouts and 25-79 min for females that called only in single bout. Similarly, intermittent calling was observed in mango fruit borer, Deanolis albizonalis (Sujatha et al., 2002) and Leucinodes orbonalis (Renuka et al., 2000). The earliest of a female initiated calling was at 04.20 h (i.e., 22.20 h) after the start of scotophase and termination of calling was found as late as 10 min after the onset of photophase. Peak calling was observed 3 h prior to end of scotophase.
The pheromone response of H. duplifascialis with wind tunnel experiment revealed that the attraction started 4-5 h. after sun set. The attraction was to the extent of 2.83-10.00 males with air current and 1.34-6.17 males without air current, using females in the bait chamber (Table 2). The virgin females attracted the males obviously by the air-borne pheromone. In the absence of air, a significant decrease in the captured numbers was recognized as the time progressed. In the experiment with air current, the air ventilation in one direction continuously forced out the pheromone, so the males could easily locate the pheromone source. Immediately after the start of ventilation, the males, which had been flying ad lib, began to orient towards the mouth of the mylar film cylinder and showed behavioural response of swift antennal movement and fluttering of wings. Usually after several unsuccessful attempts, the male entered the cylinder to reach at the opening of the opposite chamber from which the pheromone of virgin females was dispersed. The attraction was at the highest to two day old females and the lowest to freshly emerged females (<24 h); the extent being 66.67 and 18.89%, respectively, with air current and 43.33 and 8.89%, respectively, without current. In Utethesia ornatrix females, pumping frequency of ovipositor increased with increase in wind speed over a range of velocities upto 1.2 ms-1 (Conner et al., 1985).
When the male are kept in the bait chamber, the attraction of female was 7.78-12.22 and 5.56-10.00% with and without air current, respectively. Twenty females (5 moths each of freshly emerged, 1, 2 and 3 days old) kept in the bait chamber attracted 45.56 and 14.45% of male moths, whereas blank showed 4.45 and 3.33% response, with and without air current, respectively. These results are in conformity with the findings of Rajabaskar and Regupathy (2004) on Conogethes punctiferlais. The present study gave a clue about the pheromone source in jasmine bud worm and information on the time and age of pheromone release, which could be studied further and successfully exploited for monitoring and mass trapping in the pest management programmes of jasmine crop in the near future.
ACKNOWLEDGEMENT
The authors are grateful to M/S Syngenta India Ltd., Mumbai for the financial assistance provided for the study.
REFERENCES
- Mazomenos, B.E., M. Konestantopouious, D. Stefaneu, S. Skareas and L.C. Tzeiranakis, 2002. Female calling behaviour and male response to the synthetic sex pheromone components of Palpita unionalis (Lepidoptera: Pyralidae). IOBC WPRS Bull., 25: 1-10.
Direct Link