
C. Rebourg
E.R. Biodiversity and Environment, Case 5, Pl. V. Hugo, Universite de Provence, 13331, Marseilles Cedex 3, France
F. Petenian
E.R. Biodiversity and Environment, Case 5, Pl. V. Hugo, Universite de Provence, 13331, Marseilles Cedex 3, France
E. Cosson
E.R. Biodiversity and Environment, Case 5, Pl. V. Hugo, Universite de Provence, 13331, Marseilles Cedex 3, France
E. Faure
E.R. Biodiversity and Environment, Case 5, Pl. V. Hugo, Universite de Provence, 13331, Marseilles Cedex 3, France
Journal of Entomology
Year: 2006 | Volume: 3 | Issue: 3 | Page No.: 204-215







ABSTRACT
The number of species of the genus Parnassius, greatly outnumbers those in other genera of the subfamily Parnassiinae, suggests a unique evolutionary history for the Apollo butterflies. Recent extensive molecular analyses evidenced a relatively rapid radiation of Parnassius but additive studies are required for a more comprehensive understanding of the evolution of this genus. Then, the relationships between Parnassius phylogeny and host-plant preferences have been investigated using molecular phylogenetic analyses. Parnassius molecular phylogeny using a combined data set of four mitochondrial genes is congruent with the larval host-plants phylogeny inferred from chloroplast matK gene. Phylogenetic analyses evidence that Parnassius species have been divided in two groups, the Parnassius subgenus which feed mainly on Crassulaceae and all the other subgenera, containing the most basal species, which feed mainly on Corydalis. However, these co-analyses alone fail to explain all the observed patterns, indeed if host-plant shifts could explain sympatric speciations, other events, including ecological changes (habitat and elevation) have also play an important role. In conclusion, according us, only a combination of sympatric and allopatric speciations due to numerous factors could explain the Parnassius rapid adaptive radiations.
PDF Abstract XML References
How to cite this article
C. Rebourg, F. Petenian, E. Cosson and E. Faure, 2006. Patterns of Speciation and Adaptive Radiation in Parnassius Butterflies. Journal of Entomology, 3: 204-215.
DOI: 10.3923/je.2006.204.215
URL: https://scialert.net/abstract/?doi=je.2006.204.215
DOI: 10.3923/je.2006.204.215
URL: https://scialert.net/abstract/?doi=je.2006.204.215
INTRODUCTION
Swallowtail butterflies of the genus Parnassius Latreille (1804) are currently used as model organisms in several research areas, including genetics, phylogenetics of host plant utilization and mimicry, mechanisms of speciation and conservation. They also attract both museum taxonomists and amateur collectors because of their geographic variability and rarity of some species that occur in the remote alpine areas of the Himalayas, Central Asia, Tibet and other parts of northern Eurasia (in paleartic mountains). To date, the phylogenetic relationship between Parnassius and its related genera remains controversial. According to traditional studies, the genus Parnassius is classified with two other genera of similar morphology, Archon and Hypermnestra, into the tribe Parnassiini of the subfamily Parnassiinae (Munroe, 1961; Hancock, 1983). Together with the tribe Zerynthiini comprising five genera (Zerynthia, Allancastria, Sericinus, Bhutanitis and Luehdorfia), they are usually regarded as relatively primitive groups in the family Papilionidae (Munroe, 1961; Ackery, 1975; Igarashi, 1979; Hancock, 1983; Häuser, 1993). However, in recent extensive phylogenetic analyses (Omoto et al., 2004; Katoh et al., 2005) Parnassius was found to be most closely related to Hypermnestra helios, whereas Archon apollinus was more closely related to members of the tribe Zerynthiini. In addition, if previous studies have called into the question the monophyly of Parnassiinae (Häuser, 1993; Yagi et al., 1999; Caterino et al., 2001), recent molecular investigations have evidenced that the genus Parnassius constitutes a monophyletic group among the subfamily Parnassiinae, with a number of separate lineages that probably arose through a relatively rapid radiation event during evolution (Omoto et al., 2004; Katoh et al., 2005). Other genera in the same subfamily are clearly separated from Parnassius, suggesting that these genera represent evolutionary much older lineages. In addition, the number of species of the genus Parnassius which is greatly outnumbers those in other genera of the subfamily Parnassiinae, suggesting a unique evolutionary history for the Parnassius genus (~50 species on 69 divide in 8 genera). Within the genus Parnassius up to 10 species-groups or subgenera have been proposed based on morphological and behavioural studies (Bryk, 1935; Eisner, 1958, 1968; Munroe, 1961; Ackery, 1975; Hancock, 1983; Weiss, 1992, 1999). The exact number of species, however, is not known because of disagreements as to the species/subspecies rank. The morphological characteristics primarily used in the classification of species and species groups include wing pattern, venation, male genitalia, fore-tibial epiphysis and sphragis, i.e., the attachment to the end of the female abdomen made by the male secretion during copulation (Hancock, 1983). On the basis of morphological comparisons, it has been suggested that butterflies of the genus Parnassius had undergone much speciation in the Tibetan region since early Tertiary period (Hancock, 1983). However, the biological relationships among species and species-groups, as well as the evolutionary scenario of Parnassius butterflies have thus far only been speculative. In addition, despite extensive molecular analyses, the phylogenetic relationships among Parnassius sub-groups were not clear (Omoto et al., 2004; Katoh et al., 2005). Therefore, complimentary studies are required to elucidate the relationships within this genus. The number of species of the genus Parnassius (more than 70% of the subfamily), suggesting rapid adaptive radiations added to the host-plants shifts is ideal for co-analysis of animal and plant phylogenies. Indeed, it is well known that adaptation to alternate host plants can cause reproductive isolation and speciation (Dre`s and Mallet, 2002). In this study, relationships between Parnassius phylogeny using a combined data set of four mitochondrial genes and host-plant phylogeny inferred from chloroplast matK gene have been investigated.
Materials and Methods
Butterfly Taxon Sampling
The species of Parnassiinae butterflies and host-plants investigated in this study are listed in Table 1. Our choice of butterfly specimens to be analyzed was guided by the necessity to include at least one species of the major potential subgenera of the Parnassius genus (Table 1). They have been collected in broad areas of Eurasia.
DNA Extraction, PCR Amplification and Sequencing
All the chloroplast matK nucleotidic sequences of host-plants were extracted from GenBank. For the Parnassius analyses, DNA was extracted from legs or heads of dried adult butterfly according to a method described by Monte-Alegre et al. (2005). Three regions of the mitochondrial genome were amplified by Polymerase Chain Reaction (PCR) using the following oligonucleotide primers: 5’-CGCCTGTTTATCAAAAACAT and 5’-CCGGTTTGAGCTCAGATCA for the LSU(16S rRNA) gene (Simon et al., 1994); 5’-CGTAAAGTCCTAGGTTATATTCAGATTCG and 5’-ATCAAAAGGAGCTCGATTAGTTTC for the ND1 gene (Aubert et al., 1996) and 5’-GGTCAACAAATCATAAAGATATTGG and 5’-TAAACTTCAGGGTGACCAAAAAATCA for the COI gene (Folmer et al., 1994).
| Table 1: | Life history characters for Parnassius species and GenBank accession numbers for butterflies and host-plants analyzed in this study |
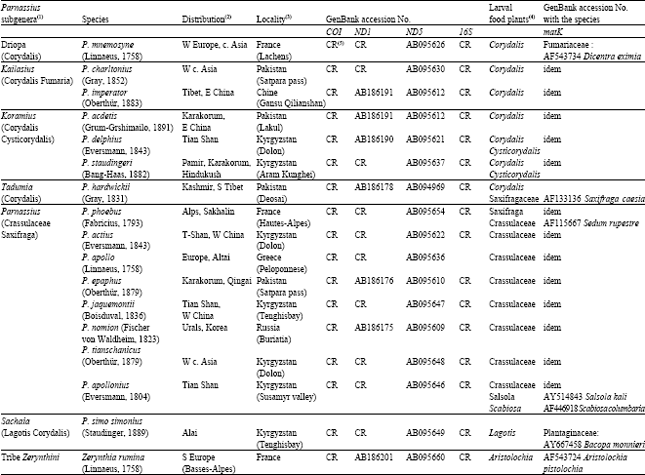 | |
| (1)With the commonly used host plants in parentheses. (2)Geographical ranges are abbreviated as follows: c., central; E, East; S, South; W, West. (3)Localities only for the samples sequenced in our laboratory. (4)Sources: (Seppänen, 1970; D’Abrera, 1990; Chou, 1994; Tuzov et al., 1997; Tolman and Lewington, 1997). (5)CR: Sequences submitted to GenBank. | |
PCR components per 50 μL reaction were as follows 50 ng template DNA, 0.2 μM of each primer, 2.0 U. HiTaq Taq polymerase, 0.2 mM dNTPs, 5 μL of the reaction buffer provided by the Taq manufacturer (Bioprobe, France). The cycling parameters were as follows 92°C for 2 min, 5 times (92°C for 40 sec 43°C for 45 sec and 62°C for 1.5 min), 30 times (92°C for 40 sec 47°C for 45 sec and 62°C for 1.5 min) and 62°C for 8 min. Using the single-stranded DNA as a template, the nucleotide sequence was determined with an automated DNA sequencer (Genome Express, Grenoble, France, http://www.genomex.com). All nucleotide sequences analyzed in this paper have been deposited in the GenBank. In addition, some Parnassiinae sequences have been extracted from GenBank (all the ND5 gene sequences, the four mitochondrial genes of the outgroups and due to problems with amplifying, seven ND1 gene sequences of Parnassius).
Phylogenetic Analyses
Sequences were aligned using Clustal W software (Thompson et al., 1994). Phylogenetic analyses were performed in PHYLIP version 3.6 alpha 3 (Felsenstein, 2002) accessed at http://www.infobiogen.fr/. Three different approaches have been used : 1) the Neighbor-Joining (NJ) method (Saitou and Nei, 1987); 2) a cladistic approach using the Maximum Parsimony (MP) criterion (Swofford, 1998); 3) Maximum Likelihood (MLH) reconstruction was performed using estimations of all model parameters from the data set. Robustness of nodes was estimated by running a bootstrap test with 100 replicates for NJ, MP and MLH trees.
RESULTS
Parnassius Genus Phylogenetic Analysis
In the LSU region, alignment gaps were observed at several nucleotide sites. Therefore, these sites were excluded and the remaining 488 sites were used for further analyses. In contrast, no alignment gaps were observed in the three protein coding genes (COI, NDI and ND5), such that the consecutive 615 sites, 455 sites and 725 sites could be used, respectively for further analyses. In order to check that there was no bias introduced by one of genes, all the partial gene sequences were analysed separately using three phylogenetic methods (unweighted Maximum Parsimony (MP), Neighbor Joining (NJ) and Maximum likehood (MLH)). In addition, in these various preliminary tests, numerous Parnassiinae sequences (Battus philenor, Iphlictides podalirius, Papilio machaon, Zerynthia rumina) available from GenBank were selected in turn or together as outgroups. In all trees obtained with sequences of these four species, the Parnassius genus formed a monophyletic group. Using a largest dataset and, respectively ND5 genes alone or both ND5 and LSU sequences, Omoto et al. (2004) and Katoh et al. (2005) have already evidenced the monophyly of this genus. In addition, alignments that contained any single outgroup taxon, or a combination of any four outgroup taxa, did not generate great variation of tree topology among the ingroup taxa for any of the algorithms that produced unrooted trees. In preliminary studies, each gene has been used separately. All the molecular phylogenetic trees obtained by the three different methods show essentially the same branching pattern and in some portions of the tree where results were varied for different methods, the bootstrap values were low (data not shown). As there was not serious incompatibility of datasets generated from the four DNA sections, we carried out a combined analysis in which 68 sequences (17 for each gene) were used, the four alignments (one by DNA partial genes) were aligned together (final alignment is available from the authors upon request).
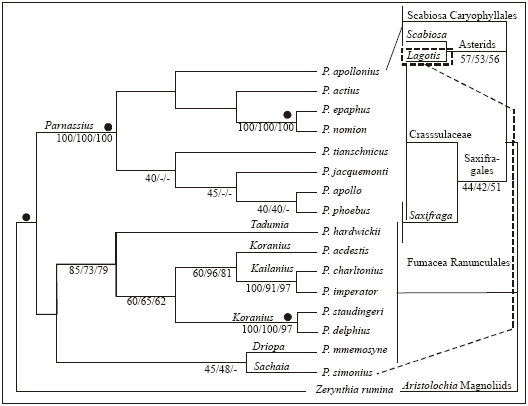 | |
| Fig. 1: | Comparison of phylogeny of Parnassius species with their host-plants phylogeny. Parnassius phylogeny has been inferred from concatenated sequences (COI, ND1, ND5 and LSU), partial matK genes has been used for plant phylogeny. As the trees are not significantly different in topology so only the Maximum Parsimony trees are shown, the branch lengths are approximate. Bootstrap values (100 replicates) are shown below branches, Maximum Parsimony (left bootstrap value), Neighbor Joining (middle bootstrap value) and Maximum likelihood (right bootstrap value), only selected values over 39 are given. In the Parnassius tree, a black circle denotes that this node has been found when the four data sets have been analysed separately, an asterisk (*) denotes that a similar topology has been found in the Omoto et al. (2004) analysis |
Rooting on Z. rumina species, the three tree-making methods (MP, NJ and MLH) produced similar topologies, which displayed one weakly supported group and one strongly supported clade corresponding to the Parnassius subgenus (Fig. 1). In the first group, two subgroups were apparent, one of them containing two species belonging to two different subgenera (Driopa and Sachaia) was not statistically supported and this branching patterns was unstable, the last one contained species belonging to three subgenera (Tadumia, Koramius and Kailasius) and was well supported by bootstrap analyses (BP = 85/73/79). In this last clade, Tadumia emerged as a sister taxon to the two other subgenera and Koramius was paraphyletic but this was not supported statistically.
Interestingly, the higher bootstrap values were found in clades which exhibited similarly high bootstrap values in phylogenetic analyses inferred by one of the four genes alone (Parnassius subgenus, P. epaphus + P. nomion and P. staudingeri + P. delphius), in addition, the phylogenetic analyses of Omoto et al. (2004), in spite of the fact that their datasets were more complete, exhibited similar groupings. In addition, other groupings which were in agreement with this previously published study are shown in Fig. 1. In addition, similarly to Omoto et al. (2004) and in spite of the fact that our data set contains only two species - but one by sub-group according to Wiess (1999) - our analyses confirm the monophyly of the Kailasius subgenus. This taxon is composed of seven species with wide distribution ranging from Central Asia through the Great Himalayas to western China. They are mostly big and brilliantly colored butterflies and usually classified under two different species-groups morphologically distinguished, viz., the Charltonius group and the Imperator group. Moreover, our analyses also evidenced that P. acdestis, which belongs to Koramius subgenera, is found rather to belong to this cluster or at least to be a sister group. In the molecular analyses of Omoto et al. (2004) and Katoh et al. (2005), P. acdestis was also found to belong to the Kailasius cluster.
Host-plants Phylogenetic Analysis
Parnassius species of our representative data set make use together in the wild of no less than seven different plant families that belong to several of the major clades of angiosperms. All the Parnassius host-plant species have not been sequenced, when no sequence of the corresponding genus has been found in GenBank, homologous sequence of the nearest taxon has been chosen. Concerning the host-plants phylogeny, the matK gene has been chosen because it has been evidenced that this gene provides good resolution within many angiosperm sub-taxa (Hilu et al., 2003). Our phylogenetic analysis of 914 unambiguous sites (Fig. 1), of which sampling is too weak, does not claim to reflect the phylogeny of magnoliophyta, however, its topology is not in conflict with other molecular analyses. Numerous molecular studies have evidenced that both magnoliids and eudicots are resolved as monophyletic lineages (Borsch et al., 2005; Löhne and Borsch, 2005). Within eudicots, which comprise about 75% of angiosperm species diversity, many phylogenetic questions persist (e.g., Soltis et al., 2000; Judd and Olmstead, 2004), for example, the position of ranunculales (Fumacea in our analysis) could differ, being either basal in eudicots or sister to a eudicot clade. In addition, if the monophyly of the core eudicot clade is not ambiguous, relationships among the major clades of this last taxon (three lineages in our dataset : saxifragales, caryophyllales and asterids) remain uncertain (Hilu et al., 2003).
DISCUSSION
Relationships Between Parnassius Phylogeny and Host-plant Utilization
In the ongoing debate about the extent of congruence to be expected between the phylogenies of insects and their host plants, butterflies of the Papilionidae family have occupied a central place ever since 1964, when Ehrlich and Raven (1964) published their seminal paper on the proposed coevolution of butterflies and plants. If only for this reason, there have been continuing and unusual efforts to collect as much information as possible on host plant utilization by swallowtail larvae.
Butterflies of the genus Parnassius are known to utilize host plants belonging to eight families, although most host-plant records are from two families (Seppänen, 1970; D’Abrera, 1990; Chou, 1994; Tuzov et al., 1997; Tolman and Lewington, 1997). Table 1 underlines that Parnassius are associated principally with two groups of plants: Fumariaceae-mainly Corydalis-and Crassulaceae-Rhodiola, Sedum, Sempervivum and related genera. In our phylogenetic analysis, the Parnassius species have been divided in two groups of which the commonly used host-plants are different, the Saxifragales (mainly Crassulaceae) for the Parnassius subgenus and Fumariaceae and/or Lagotis for the other subgenera. Parnassius species which feed on a high altitude Scrophulariaceae (Lagotis) constitute a localized and interesting exception concerning only species of the Simo group which belongs to the Sachaia subgenus (Weiss, 1999).
Interestingly, in our data set, two species which emerged as sister group to many others (P. apollonius/other Parnassius species of this subgenus and P. harwickii/species belonging to Koramius and Kailasius subgenera) have larvae which could consume host-plants belonging to two different plant orders. In their NJ tree based on the ND5 sequence, Omoto et al. (2004) have evidenced similar topologies, P. apollonius (with P. honrathi) is the sister group of the other species of the Parnassius subgenus and P. harwickii is the sister group of all the Parnassius species except them belonging to the Parnassius subgenus. However, these topologies in the two studies have not supported statistically.
The Ancestral Parnassius Host-plant and the Various Shifts
Is it possible to guess at ancestral host plant utilization in Parnassius species? Obviously, the original food plants of the Parnassiinae-Zerynthiinae phylum are the Aristolochiaceae, still used by Archon and all Zerynthiinae. The sister genus of Parnassius, Hypermnestra, lives on Zygophyllaceae plants (Zygophyllum, Halimiphyllum) from the arid regions of western central Asia (D'Abrera, 1990; Tuzov et al., 1997). It is thus likely that the stem common to both lines had already left the Aristolochiaceae. Interestingly, Parnassius glacialis Butler, 1866 (Driopa subgenus, mnemosyne group) whose principal host-plants are Corydalis still feed on Aristochia (Chou, 1994). This parnassian species inhabits arctic or mountainous area of the Northern Hemisphere and has morphological characteristics that were suggested to be primitive (Nemoto and Inomata, 1994; Weiss, 1999).
The most basal species of Parnassius in the phylogenetic trees of the extensive studies of Omoto et al. (2004) and Katoh et al. (2005) are linked to Corydalis, a genus which contains about 320 species, mainly localized in central Asia (Heywood, 2005). Two other elements are in favour of this hypothesis, in one hand, most of the species of five on six subgenera feed mainly on plants of this genus and in another hand, the feed choice of P. glacialis evidenced probably the first ancestral plant shift.
Inside of Parnassius, the major foodplant change concerns the apollo group (Parnassius subgenus), which have adopted another group of plants adapted to mountains, the Crassulaceae. These host plants are a morphologically diverse and systematically complex angiosperm family comprising 35 genera and 1500 species (Berger, 1930). The family inhabits primarily semiarid and mountainous habitats and even if now the distribution is nearly cosmopolitan, most of the species still exhibit xerophytic adaptations (e.g., succulent leaves, a thick waxy cuticle and crassulacean acid metabolism). Therefore, the second major change has been performed inside of the same habitat allowing sympatric speciations.
The same is observed with a more restricted shift, that from Corydalis to Lagotis in the simo group of the subgenus Sachaia-the other species of this subgenus remains linked to Corydalis (Korshunov and Gorbunov, 1995): Lagotis lives on high elevation screes, where certain species of Corydalis happen to be numerous. This is true also for the shift of phoebus to the widespread Saxifraga like S. aizoides in the Alps: this plant lives in exactly the same habitat as the supposedly genuine foodplant, Rhodiola rosea, the shores of torrents. Interestingly, this last shift within species which belong to the same plant order (saxifragales) and certain populations of the species still feed on Crassulaceae like Sedum or Rhodiola species.
Is Chemistry Could Unit All the Parnassius Host-plants
A number of aposematic butterfly sequester unpalatable or toxic substances from their host plants rather than manufacturing their own defensive substances (Nishida, 2002). One type of toxic compounds among the most used by butterflies is alkaloid. Numerous alkaloids have been isolated in species belonging to all the Parnassius host-plants genera (Willaman and Schubert, 1961; Israilov et al., 1984; Zhu, 1998; Seger et al., 2004; Zhao and Ding, 2004; Endonova and Antsupova, 2005; Li et al., 2005) except for Crassulaceae. However, if in this family, all the genera have not been chemically investigated and these toxic compounds have been found at least in Sedum and Rhodiola species (Steven et al., 1995; Tolonen, 2003).
The Corydalis genus is closely related to the Papaveraceae, they are toxic and contain numerous alkaloïds. Parnassius have a wing pattern suggestive of aposematism and they display their eyespots with stridulation when disturbed at rest (Descimon, 1965); the larvae can be mimetic of toxic millipedes (Scott, 1986; Deschamps-Cottin and Descimon, 1996). It is therefore likely that Parnassius store toxic compounds produced by Corydalis.
Crassulaceae, the other preferred host plants, also contain alkaloids and can provide protective compounds as well, for example, piperidine alkaloids are widely distributed in Sedum species (Hegnauer, 1989). In addition, it is known that sarmentosin, a bitter-tasting cyanoglycoside produced by Crassulaceae that is not cyanogenic itself, occurs in Parnassius butterflies (Nishida and Rothschild, 1995). These cyanophilic insects apparently show host-exploitation patterns based on innate resistance to the toxins (Nishida, 2002).
The picture of Parnassius host-plants is somewhat complicated by the fact that data concerning their chemical characteristics are very partial, for example, it would be interesting to know if a (secondary) chemical product, if it exists, units the Parnassius host-plant families. We hypothesise that further extensive chemical investigations could give interesting results. For example, all the Euphydryas host-plants, which belong to 13 different families are united by the presence of a group of secondary defense chemicals known as iridoids (Jensen et al., 1975). Both are known to be extremely and Bowers and Puttick (1986) have shown that iridoid glycosides are necessary feeding stimulants for Nearctic Euphydryas larvae and that the larvae are able to sequester these compounds for use in their own chemical defense (Zimmermannn et al., 2000). Euphydryas larvae are considered aposematic and they are unpalatable to birds (Bowers 1981). Interestingly, iridoid glucosides have been found in Lagotis (Yang et al., 2004) and in Crassulaceae (Tolonen, 2003) and a putative precursor of seco-iridoid has been detected in Scabiosa (Horn et al., 2001). However, to date, it is not known, if some Parnassius species were able to sequester these compounds.
In addition, chemoreception is an essential function in the recognition of host plants for phytophagous insects. Detection of a specific repertoire of plant secondary metabolites by olfaction or contact chemoreception plays an important role in determining whether or not a plant is suitable (Nishida, 1995; Ono and Yoshikawa, 2004). And we cannot exclude phytochemical mediators serving as attractants, repellents, stimulants, or deterrents in oviposition behavior of Parnassius could units their host-plants.
Important Role of Ecological Factors (Habitat and Elevation) in Parnassius Evolution
According to Hancock (1983) the center of diversity of the Parnassius is in the Central Asian high plateau and the time of first divergence. The adaptation to high elevation habitats seems to have occurred at the second step of Parnassius evolution : the most basal branch (Dryopa) is, in its generality, associated with low mountain, semi-forested biomass. In central Asia, habitats from mountain zone clearings to the highest screes, ridges and slopes up to snow line are colonized by at least one species of Parnassius (and in Ladakh, up to seven species can be observed around Nimaling, 4000-5500 m : Weiss, 1992). This adaptation to higher elevation could explain that Glacial periods are likely to have been favourable to the extension of Parnassius; hence the wide distribution of the P. Apollo species, which reached the mountains of Andalucia as well as the Etna in Sicily and Taygetos mountain in southern Greece.
CONCLUSIONS
Judging from the branching pattern of their phylogenetic trees, which exhibit that the Parnassius clades appeared to be connected to each other by relatively short internal branches, Omoto et al. (2004) and Katoh et al. (2005) have suggested that the genus Parnassius would seem to have originated by a relatively rapid radiation. According to these authors, the time of the rapid radiation took place sometime in the late Tertiary period (20-30 MYA BP). It has been proposed (Mitter and Farrell, 1991) that the diversity of plants, in particular their chemical diversity, could be a factor involved in the strong diversification of herbivorous insects. According to what could be called the ‘explosive adaptive radiation theory’, a ‘key’ character evolves in a lineage that enables it to explore new niches. The availability of new resources as well as new areas to colonize, with other selection pressures and release from competition, could promote speciation processes (Futuyma, 1986). As soon as an insect has been able to overcome chemical defenses of plants, an opportunity for diversification can occur (Mitter et al., 1988). One of the parsimonious strategies for the phytophagous insects is shifts within plants which produced similar secondary toxic compounds, for example, the evolutionary history of host-plant use in the tribe Melitaeini evidenced that plant chemistry is a more conservative trait than plant taxonomy (Wahlberg, 2001). However, the host-plants shifts alone could not explain the relatively rapid radiation event during Parnassius evolution. Some ecological traits as habitat and elevational changes are probably also play an important role. According us, only a combination of sympatric and allopatric speciations due to numerous factors could be explain these rapid radiation events. This study evidence the two future areas of research which are required for a more comprehensive understanding of Parnassius evolution: first, extensive chemical investigations of the Parnassius host-plants; second, analyses of all the ecological factors in a historical perspective.
ACKNOWLEDGMENTS
The authors are most grateful to Henri Descimon for supplying the butterfly samples.
REFERENCES
- Aubert, J., B. Barascud, H. Descimon and F. Michel, 1996. Systematique moleculaire des Argynnes (Lepidoptera: Nymphalidae). CR Acad. Sci. III-Vie, 319: 647-652.
Direct Link - Caterino, M.S., R.D. Reed, M.M. Kuo and F.A.H. Sperling, 2001. A partitioned likelihood analysis of swallowtail butterfly phylogeny (Lepidoptera: Papilionidae). Syst. Biol., 50: 106-127.
PubMedDirect Link - Dres, M. and J. Mallet, 2002. Host races in plant-feeding insects and their importance in sympatric speciation. Philos. Trans. Roy. Soc. Lond. B, 357: 471-492.
CrossRefDirect Link - Ehrlich, P.R. and P.H. Raven, 1964. Butterflies and plants: A study in coevolution. Evolution, 18: 586-608.
Direct Link - Endonova, G.B. and T.P. Antsupova, 2005. Dynamics of accumulation of alkaloids and flavonoids in Scabiosa comosa Fisch. ex Roem. et Schult. and Scabiosa ochroleuca L. Russ. J. Ecol., 36: 48-51.
CrossRefDirect Link - Folmer, O., M. Black, W. Hoeh, R. Lutz and R. Vrijenhoek, 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol., 3: 294-299.
PubMedDirect Link - Hilu, K.W., T. Borsch, K. Muller, D.E. Soltis and P.S. Soltis et al., 2003. Angiosperm phylogeny based on matK sequence information. Am. J. Bot., 90: 1758-1776.
Direct Link - Horn, M.M., S.E. Drewes N.J. Brown, O.Q. Munro, J.J. Meyer and A.D. Mathekga, 2001. Transformation of naturally-occurring 1,9-trans-9,5-cis sweroside to all trans sweroside during acetylation of sweroside aglycone. Phytochemistry, 57: 51-56.
CrossRefDirect Link - Judd, W.S. and R.G. Olmstead, 2004. A survey of tricolpate (eudicot) phylogenetic relationships. Am. J. Bot., 91: 1627-1644.
Direct Link - Katoh, T., A. Chichvarkhin, T. Yagi and K. Omoto, 2005. Phylogeny and evolution of butterflies of the genus Parnassius: Inferences from mitochondrial 16S and ND1 sequences. Zool. Sci., 22: 343-351.
Direct Link - Lohne, C. and T. Borsch, 2005. Molecular evolution and phylogenetic utility of the petD group II intron: A case study in basal angiosperms. Mol. Biol. Evol., 22: 317-332.
CrossRefDirect Link - Mitter, C., B. Farrel and B. Wiegmann, 1988. The phylogenetic study of adaptive zones: has phytophagy promoted insect diversification? Am. Natl., 132: 107-128.
Direct Link - Monte-alegre A., Z. Rakotomalala, M. Leveugle, F. Coulier and E. Faure, 2005. Ecological, vertical and elevational distributions of Heteropsis (Lepidoptera, Satyrinae) in the rainforest of Masoala (NE Madagascar). J. Zool. Syst. Evol. Res., 43: 214-222.
CrossRefDirect Link - Nishida, R. and M. Rothschild, 1995. A cyanoglucoside stored by a Sedum-feeding Apollo butterfly, Parnassius phoebus. Experientia, 51: 267-269.
CrossRefDirect Link - Nishida, R., 2002. Sequestration of defensive substances from plants by Lepidoptera. Ann. Rev. Entomol., 47: 57-92.
CrossRefDirect Link - Omoto, K., T. Katoh, A. Chichvarkhin and T. Yagi, 2004. Molecular systemtics and evolution of the Apollo butterflies of the genus Parnassius (Lepidoptera: Papilionidae) based on mitochondrial DNA sequence data. Gene, 326: 141-147.
CrossRefDirect Link - Ono, H. and H. Yoshikawa, 2004. Identification of amine receptors from a swallowtail butterfly, Papilio xuthus L. Cloning and mRNA localization in foreleg chemosensory organ for recognition of host plants. Insect. Biochem. Mol., 34: 1247-1256.
Direct Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Seger, C., S. Sturm, E.M. Strasser, E. Ellmerer and H. Stuppner, 2004. 1H and 13C NMR signal assignment of benzylisoquinoline alkaloids from Fumaria officinalis L. (Papaveraceae). Magn. Reson. Chem., 42: 882-886.
PubMedDirect Link - Simon, C., F. Frati, A. Beckenbach, B. Crespi, H. Liu and P. Flook, 1994. Evolution, weighting and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am., 87: 651-701.
Direct Link - Soltis, D.E., P.S. Soltis, M.W. Chase, M.E. Mort and D.C. Albach et al., 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL and atpB sequences. Bot. J. Linn. Soc., 133: 381-461.
Direct Link - Stevens, J.F., H. Hart, R.C.H.J. Van Ham, E.T. Elema, M.M.V.X. Van Den Ent, M. Wildeboer and J.H. Zwaving, 1995. Distribution of Alkaloids and Tannins in the Crassulaceae. Biochem. Syst. Ecol., 23: 157-165.
CrossRefDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Wahlberg, N., 2001. The phylogenetics and biochemistry of host-plant specialization in Melitaeine butterflies (Lepidoptera: Nymphalidae). Evolution, 55: 522-537.
PubMedDirect Link - Yagi, T., G. Sasaki and H. Takebe, 1999. Phylogeny of Japanese Papilionid butterflies inferred from nucleotide sequences of the mitochondrial ND5 gene. J. Mol. Evol., 48: 42-48.
CrossRefDirect Link - Yang, X.D., J.F. Zhao, S.X. Mei, J.X. Pu, H.B. Zhang and L. Li, 2004. Two new iridoid glucosides from Lagotis yunnanensis. Chin. Chem. Lett., 15: 49-51.
Direct Link - Zhao, Y.X. and X.B. Ding, 2004. Studies on the alkaloids from Salsola collina Pall. Yao. Xue. Xue. Bao., 39: 598-600.
Direct Link - Zimmermann, M., N. Wahlberg and H. Descimon, 2000. Phylogeny of Euphydryas checkerspot butterflies (Lepidoptera: Nymphalidae) based on mitochondrial DNA sequence data. Ann. Entomol. Soc. Am., 93: 347-355.
Direct Link