
A. N. Mahar
Sindh Agriculture University, TandoJam, Pakistan
D. A. Darban
Sindh Agriculture University, TandoJam, Pakistan
A. G. Lanjar
Sindh Agriculture University, TandoJam, Pakistan
M. Munir N.D.
School of Plant Sciences, University of Reading, Reading, UK
Jan N.G.M Hague
School of Plant Sciences, University of Reading, Reading, UK
S. R. Gowen
3Department of Agriculture, University of Reading, UK
Journal of Entomology
Year: 2005 | Volume: 2 | Issue: 1 | Page No.: 92-96







ABSTRACT
Susceptibility of late instar vine weevil Otiorhynchus sulcatus larvae and pupae to four species entomopathogenic nematodes were tested. Bioassays on production and infectivity to larvae and pupae were compared for two steinernematids and two heterorhabditis such as Steinernema carpocapsae, S. feltiae, Heterorhabditis indica and H. bacteriophora. Nematodes production of all species was determined by the number infective juveniles (IJs) established in vine weevil larvae and pupae O. sulcatus using sand and filter paper bioassay. S. feltiae produced the maximum number in larvae and pupae at 20°C as compared to other nematodes but production of H. indica, was better at 25°C in larvae and pupae followed by H. bacteriophora, S. carpocapsae and Infectivity test of larvae and pupae was also done in sand media. Infective juveniles recovered from larvae and pupae when infected with S. feltiae produced maximum infective juveniles at 20°C temperatures than all other isolates. H. bacteriophora produced higher number of IJs in larvae and pupae than all other nematode isolates at 25°C. This paper indicates the application of nematodes with the knowledge of insect pest biology represents a possible new strategy for O. sulcatus larvae and pupae.
PDF Abstract XML References
How to cite this article
A. N. Mahar, D. A. Darban, A. G. Lanjar, M. Munir N.D., Jan N.G.M Hague and S. R. Gowen, 2005. Influence of Temperature on the Production and Infectivity of Entomopathogenic Nematodes Against Larvae and Pupae of Vine Weevil, Otiorhynchus sulcatus (Coleoptera:Curculionidae). Journal of Entomology, 2: 92-96.
DOI: 10.3923/je.2005.92.96
URL: https://scialert.net/abstract/?doi=je.2005.92.96
DOI: 10.3923/je.2005.92.96
URL: https://scialert.net/abstract/?doi=je.2005.92.96
INTRODUCTION
Black vine weevil, Otiorhynchus sulcatus, (F.) Coleoptera:Curculionidae is an important pest of many economic important plants mostly strawberries, raspberries and rhododendrons. The larvae and adults attack on the roots of many annual perennial ornamental and soft fruit plants. Young larvae or grubs are creamy white or dirty white to brown, have no legs and feed on fine roots and crowns in mid-summer, or over winter in the soil and cause heaviest damage in the spring. Black vine weevil O. sulcatus pupae are soft and white. Adults emerge after a short pupation period in April and May. There is only one generation per year[1]. Adults of this insect pest eat notches in the leaves, feed on foliage but most damage to roots is caused by the late instars of larvae. Plants become stunted and darkened and this damage can weaken or kill the plant. Injured plants have a stunted appearance; the leaves are closely bunched and are dark and blue-green. The fine roots have been destroyed and some times even the hard fibrous roots have been eaten.
Entomopathogenic nematodes, the Steinernematids and Heterorhabditids have been utilized as the biological control agents for arrange of pests throughout the world. They are potential alternatives to chemical control of many insect pests. However there is considerable variation in the production and infectivity of entomopathogenic nematodes and no single species or strain is suitable for controlling all or even most insect species[2]. Entomopathogenic nematode have emerged as excellent candidates for biological control of insect pests. They attributes making the nematodes ideal biological insecticides include their broad host range, high virulence, safety for non-target organisms and high efficacy in favourable habitats. Progress achieved in liquid fermentation, formulation stability and application strategy has allowed nematode-based products to become competitive with chemical insecticides in medium and high valued crops on the basis of cost/ benefit ratio and ease of application[3]. Evaluation of the potential entomopathogenic nematodes for use as a biological control agent against a selected target insect pest usually includes an assessment of effectiveness in relation to all developmental stages of the pest. Entomopathogenic nematodes enter the host through natural body openings[4]. Spiracular orifices in pupal cases are an obvious potential point of entry[5]. However, pupae of different insect species vary in susceptibility to nematode infection[6].
Temperature is known to influence the efficacy of these nematodes as biological control agents. Extremes in temperatures affect both the target insect and the nematode parasites. However, most studies appear to regard only the nematode as important. Temperature affects reproduction capability nematode growth and development, Infection potential of IJs and emergence time, migration activity, sex ratios and insect death rates[5]. Infectivity of entomopathogenic nematodes depends also on the physiological stage of their infective juveniles. As they are non feeding, IJs use their stored lipids as a source of energy[7].
Knowledge of the temperature limits and optima for infection of specific nematodes is of great importance to field application. However, when applying nematodes for the control of vine weevil larvae, pupae or against adults, the soil environment often acts as a buffer against temperature extremes.
The aim of this study was to examine the differences in the reproduction rate between most virulent nematodes against a serious insect pest O. sulcatus larvae and pupae which is causing a serious damage to many vegetable crops. This study was undertaken to evaluate the effectiveness of some strains of Steinernematids and Heterorhabditids against larvae and pupae of black vine weevil.
MATERIALS AND METHODS
O. sulcatus larvae were maintained on the roots of potted strawberry plants (Fragaria ananassa cv. Pegasus) on which natural population of vine weevils had oviposited. Plants were grown in the glasshouse at 20°C in the Department of Agriculture, University of Reading. Pupae were obtained from the infected soil in plant pots already in pots last instars of vine weevil larvae were found attached with the roots of plants. S. carpocapsae (All isolate) and cultured at 25°C originally obtained from by Biosys, USA, S. feltiae, (UK isolate cultured at 25°C) and H. bacteriophora (HW79 isolate cultured at 25°C) nematodes IJs suspensions were supplied by CAB Institute of Parasitology, St. Albans, UK. H. indica (Pakistan isolate cultured at 25°C) nematode was supplied by Dr. Shahina Fayyaz from Pakistan Nematological Research Center Karachi, Sindh Pakistan. Nematodes were cultured in the greater wax moth, Galleria mellonella. S. carpocapsae and S. feltiae were stored at 7°C and other two Heterorhabditis, such as H. indica and H. bacteriophora were stored at 15°C. The fresh IJs were used within one week of harvesting from white traps using the techniques described[4].
Experiment 1. The production of infective juveniles O. sulcatus larvae at different temperatures: The production of IJs of all isolates in O. sulcatus larvae was investigated at 20 and 25°C. Single last instar larvae of O. sulcatus same size and weight were infected with 100 IJs from each species of isolate in multi-well dishes with 12 cells (2.5 cm in diameter and 2.0 cm in depth) filled with eight gram of moist autoclaved sand (14% MC). In this experiment IJs were applied to each cell of muti-well dishes, were sealed with paraflim to avoid desiccation and placed in incubator at 20 and 25°C. After 5 days exposure the each larvae were transferred to separate White trap containing filter paper with distilled water and the number of emerging IJs were counted every two days until there was no further recovery. Replication was 12 fold.
Experiment 2. The production of infective juveniles in O. sulcatus pupae at different temperatures: O. sulcatus pupae were used for investigating the production of IJs at 20 and 25°C. Single pupae of same size and weight were infected with 100 IJs of each species of isolates in multi-well dishes with 12 cells (2.5 cm in diameter and 2.0 cm in depth) filled with eight gram of moist autoclaved sand (14% MC). Experimental procedure was same as done for larvae. Containers were placed in incubator at 20 and 25°C. After 5 days exposure the each pupae were transferred to separate White trap and the number of emerging IJs were counted every two days until there was no further recovery. Replication was 12 fold.
Experiment 3. Effect of temperature on infectivity of O. sulcatus larvae: Infectivity of four entomopathogenic nematodes to O. sulcatus larvae was compared at two different temperatures using same sand-based assay[2]. In this experiment last instar of O. sulcatus larvae were used. Single larvae of same age, size and weight were infected with 100 IJs of each isolate using sand media in same manner as described already in previous experiment. Multi-well dishes were incubated at 20 and 25°C. After 2 days exposure the larvae were transferred to petridishes containing Ringer solution and were dissected. Total numbers of emerging IJs were counted. Replication was 12 fold.
Experiment 4. Effect of temperature on infectivity test of O. sulcatus pupae: Single pupae of same age, size and weight of O. sulcatus were infected with 100 IJs of each isolate using sand media in same manner as described already for O. sulcatus larvae. Multi-well dishes were incubated at 20 and 25°C. After 2 days exposure the pupae were transferred to petri dishes containing Ringer solution and were dissected. Total number of emerging IJs were counted. Replication was 12 fold.
Statistical analysis: Data were analysed using techniques of GENSTST-5, Release 4.2 (Lawes Agricultural Trust, Rothamsted Experimental Station, UK). MS Excel was used for graphs.
RESULTS
Production of different juveniles in O. sulcatus larvae and pupae: The number of infective juveniles was produced from all four entomopathogenic nematodes when they were infected but there was significant difference at both temperatures (p<0.05). At 20°C maximum number (943 per larva) was produced than 25°C (852.0). The nematode production was found highly significant (p=0.001). The maximum number (1022) of IJs of H. indica established in O. sulcatus larvae at both temperatures, followed by H. bacteriophora (944) and S. feltiae (819) (Fig. 1a). Temperature X nematode interaction was found highly significant p<0.001. At 20°C, S. carpocapsae produced maximum number of IJs (1035), followed by S. feltiae (972) and H. indica (924). At 25°C the maximum number (1120) IJs of H. indica established in larvae, followed by H. bacteriophora (1047) and S. feltiae (666). Temperature showed the significant effect for establishment of nematodes (Fig. 1b).
In case of pupae production experiment temperature was found non significant. However at 25°C produced highest number of IJs (614) as compared to 20°C (577). Nematode isolate production in pupae was highly significant (p<0.001). H. indica produced the maximum number (710) followed by H. bacteriophora (609) and S. carpocapsae (538) in per pupa.
Interaction of temperature and nematode was highly significant (p<0.001). At 20°C maximum number per pupa produced by S. feltiae (730) followed by S. carpocapsae (674) and H. indica (507). At 25°C H. indica produced maximum number (912) per larva followed by H. bacteriophora and S. carpocapsae (402) (Fig. 2a and b).
Infectivity and mortality of O. sulcatus larvae and pupae: In case of infectivity test temperature was found significant (p<0.01) and maximum number (57.01) produced at 25°C as compared to 20°C (42.3) per larva. Nematode effect was non significant. However, S. feltiae produced maximum number (54.1) followed by S. carpocapsae (49.9) and H. indica (47.7). Whereas, interaction of temperature and nematode resulted non-significant, but maximum number (56.2) per larvae was penetrated by S. feltiae, most virulent and appeared to be more infective at 20°C in O. sulcatus larvae, followed by S. carpocapsae (42.8), H. indica (37) when tested in sand arenas for infectivity test (Fig. 3).
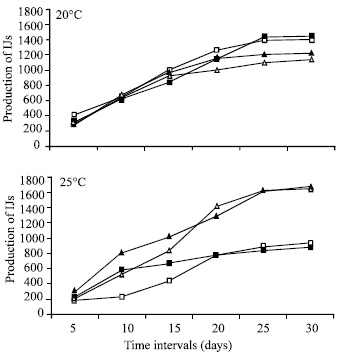 | |
| Fig. 1: | The number of infective juveniles produced by S. carpocapsae (□), S. feltiae (■), H. Indica (▲) and H. bacteriophora (Δ) in Black Vine Weevil (O. Sulcatus) larvae at 20 and 25°C (SED value: 57.5) |
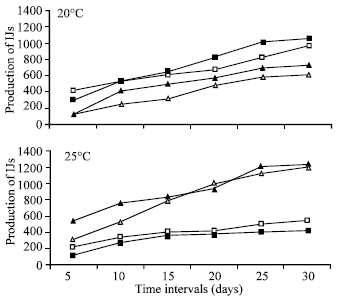 | |
| Fig. 2: | The number of infective juveniles produced by S. carpocapsae (□), S. feltiae (■), H. Indica (▲) and H. bacteriophora (Δ) in Black Vine Weevil (O. Sulcatus) pupae at 20 and 25°C (SED value: 36.8) |
Number of IJs found in O. sulcatus larvae when tested for infectivity at 25°C showed a significant increased number (61.2) of H. bacteriophora, followed by H. indica (58.4), S. carpocapsae (57) (Fig. 3).
Temperature showed significant (p<0.05) effect for infectivity test in O. sulcatus pupae. Maximum number (37.1 per pupa) produced at 25°C as compared to 20°C (29.7). Nematode effect was non-significant however maximum number (34.6 per pupae) produced by S. carpocapsae followed by S. feltiae (33.4) and H. indica (32.9) (Fig. 4).
Interaction of temperature and nematodes showed non-significant effect however maximum number (35.5 per pupa) penetrated by S. feltiae at 20°C followed by S. carpocapsae (32.2) and H. indica (26.5). At 25°C the maximum number (41) produced by H. bacteriophora followed by H. indica (39.2) and S. carpocapsae (37.0 per pupa) (Fig. 4).
DISCUSSION
Temperature tolerance of nematodes varied between nematode species. The behaviour of species and affected infection response indicates that these are capable of entering on a low or high temperature. In a sand based assay, temperature affects the infectivity (penetration ability and insect mortality) of different nematodes. The bioassays described in this study tested that insect parasitic nematode S. feltiae remained most effective at low temperature 20°C and produced the maximum number of Juveniles whereas H. indica produced more number of juveniles at higher temperature 25°C when tested for production but H. bacteriophora produced higher number of nematodes when tested for infectivity. Present findings are similar[8] who reported that both strains of S. feltiae nematodes and the LT50 decreased as the temperature increased. No significant difference was found between the LT50 of two strains at 15, 20 and 25°C. Furthermore they suggested that strains of S. feltiae could be more effective at low temperatures against Carrot weevil adults Listronotus oregonensis. Soil environment holds numerous organisms that are antagonistic to entomopathogenic nematodes including bacteria, predatory nematodes, arthropods and microorganisms[9].
Temperature has a direct influence on the activity of the nematode and on the pathogenicity of the bacterial symbionts[10]. Cold temperature causes a temporary loss in the ability of nematodes to infect a host, although they appear to move normally[11].
The present findings on the temperature preference of the nematodes for infection and development agree with the published literature. For example, S. feltiae has been reported to be infective between 4 and 25°C and infectivity has been reported to occur over a wider range of temperatures than development[12].
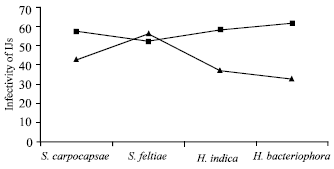 | |
| Fig. 3: | The number of infective juveniles penetrated by S. carpocapsae, S. feltiae, H. Indica and H. bacteriophora into the body of a single Black Vine Weevil (O. Sulcatus) larvae (SED value: 9.41) at 20 (▲) and 25°C (■) |
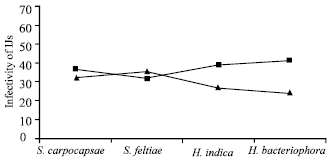 | |
| Fig. 4: | The number of infective juveniles penetrated by S. carpocapsae, S. feltiae, H. Indica and H. bacteriophora into the body of a single Black Vine Weevil (O. Sulcatus) pupae at 20 (▲) and 25°C (■) at SED value: 6.30 |
S. carpocapsae development did not occur at 10 and above 33°C, whilst at 30°C nematodes developed to adults but did not reproduce[13]. The optimal temperature for infection, production and development of Steinernema spp. lie between 15 and 20°C[14].
Similar findings were reported when three species of entomopathogenic nematodes, S. kraussei, S. feltiae and H. megidis against black vine weevil O. sulcatus small larvae, large larvae and pupae at different temperatures were investigated. Their results revealed a significant effect of temperature on O. sulcatus larvae and pupae but also showed differential levels of mortality[15].
Infectivity was determined by the number of infective juveniles established in Cylas formicarius larvae when it was indicated that there was significant difference in nematode establishment among experimental arenas for each nematode tested. S. carpocapsae strains were favoured in petri plate arenas while Heterorhabditis strains favoured in sand arenas. Further more they reported that nematode establishment increased with time for the first 4 to 8 days and then leveled off for the reaming test period for most nematode-bioassay combination[16].
Thermal niche for infection, establishment and reproduction of some entomopathogenic nematodes was reported[17]. They found that S. riobravis infected G. mellonella larvae at the widest temperature range 10-39°C. whereas S. feltiae at the narrowest (18-30°C). For reproduction S. glaseri 10-37°C, S. carpocapsae 20-30°C, S. scapterisci 20-32°C. S. riobravis 20-35°C. They were more adapted to warm temperature reproduction and S. feltiae to cooler temperature 10-25°C.
The efficacy of two Heterorhabditis isolates for control of vine weevil in both open ground and out door containers was compared. The experimental results showed that nematodes were more effective in containers than they were in open ground. It was concluded that antagonism in the soil was probably a contributing factor[18].
For controlling black vine weevil larvae and pupae in soil environment at an early stage of infestation can be possible with the knowledge of virulent biocontrol agent which has enough capability of production and efficacy against this pest. Damage to plant roots can be minimized and economic loss can be prevented.
REFERENCES
- Lindegren, L.E., K.F. Meyer, T.J. Henneberry, P.V. Vail, L.J. Forlow Jech and K.A. Valero, 1993. Susceptibility of pink bollworm (Lepidoptera: Gelechiidae) soil associated stages to the entomopathogenic nematode Steinernema carpocapsae (Rhabditida: Steinernematidae). Southwestern Entomol., 18: 113-120.