
Junior Baudoin Wouokoue Taffo
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
LiveDNA: 237.26373
Victor Francois Nguetsop
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
Grace Mendi Anjah
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
Marie Caroline Momo Solefack
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
Walter Ndam Tacham
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
Samuel Severin Kenfack Feukeng
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
Journal of Applied Sciences
Year: 2019 | Volume: 19 | Issue: 2 | Page No.: 68-76







ABSTRACT
Background and Objective: The great variety of phenological patterns in tropical mountain trees must not be explained only by climatic factors. Considering that soil water availability influences plant water status and total metabolism, soil conditions and altitude are also expected to be important for the regulation of plant reproductive and vegetative activities over time. The aim of this study was to investigate the effect of climatic factors and altitude on the phenology of Entada africana and Tephrosia vogelii in three altitudinal zones of the Bambouto Mountains, West Cameroon. Materials and Methods: Phenological (flushing, leaf fall, flowering and fruiting) data were monitored monthly in the dry season and every 2 weeks in the rainy season from October, 2012 to September, 2013 for 120 trees of two tree species (60 trees for each species and 20 in each zone) in three altitudinal zones: The lower zone (1100-1600 m), the mid-zone (1600-2000 m) and the upper zone (above 2000 m). Results: Each species phenological patterns were alike and marked by seasonality on the phenophases in the same zone. However, the frequency, peak and intensity of the phenophases in the three altitudinal zones were distinct. Leaf initiation activity in E. africana and T. vogelii were associated with maximum temperature in dry season. Flowering activity started in the rainy season. Fruit initiation were associated with rainfall in rainy season, the dehiscence of fruits was completed in dry season. Precocity of leaf-initiation in the dry season and delayed of flowering, fruiting in the rainy season in response to the effect of climatic conditions and altitude was observed in the upper zone. Conclusion: Climate and altitude were the important factors affecting phenological patterns in tropical mountain tree species.
PDF Abstract XML References Citation
Received: December 27, 2018;
Accepted: February 01, 2019;
Published: April 05, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Junior Baudoin Wouokoue Taffo, Victor Francois Nguetsop, Grace Mendi Anjah, Marie Caroline Momo Solefack, Walter Ndam Tacham and Samuel Severin Kenfack Feukeng, 2019. Phenological Behaviour of Tropical Tree Species in Three Altitudinal Zones of Bambouto Mountains, West Cameroon. Journal of Applied Sciences, 19: 68-76.
DOI: 10.3923/jas.2019.68.76
URL: https://scialert.net/abstract/?doi=jas.2019.68.76
DOI: 10.3923/jas.2019.68.76
URL: https://scialert.net/abstract/?doi=jas.2019.68.76
INTRODUCTION
Seasonal duration of leafing, flowering and fruiting mainly determine phenological behaviour in tropical trees. Plant phenology is the study of the timing of the development of vegetative and reproductive organs, most notably leafing, flowering and fruiting1. Phenological events are controlled by a combination of abiotic and biotic factors that determine their occurrence or inhibition2. In tropical regions, seasons are often marked by differences in rainfall, with life-history events occurring in response to water availability. The importance of these periodic phenomenas in plants is the quest to grab temporal, spatial and stational progress of their apparition3. The initiation of growth in plants and changes in phenology are governed by various environmental factors and the influence of temperature and moisture has been studied by Morellato et al.4 and Marques et al.5. Several authors have studied the role of soil water reverse on leafing, flowering and fruiting6,7. However, the simultaneity action of the factors does not permit to attribute water the exclusive role in their occurrence6. Hamann8 suggested that climatic factors are not directly responsible for triggering and synchronizing phenological events. Since phenological processes in plants are governed by climate, changes in global climate may influence the timing, duration and synchronization of phenological events in plant species. Therefore, a significant interest in phenological analysis of plants have emerged and is often considered as a useful tool to address critical questions related to monitoring modelling of climate change9,10.
Therefore, information regarding the phenological patterns is important for understanding the biology of plant species and also other organisms associated with them. Such studies are essential for the conservation of tree genetic resources and forestry management. Study of plant phenology constitutes a necessary tool for their amelioration. As such, they enrich the knowledge of species autecology. It also helps to understand the ecological adaptations and community level interactions of plant species.
The phenological patterns of the tree species in Cameroon concern punctual observations marked and observed during the botanic inventories11, those in the different volumes of the Cameroon flora written by several researchers under the supervision of Aubreville12 and recently Feteke et al.13 in three commercial trees of central Africa in the Eastern region of Cameroon. There are very few studies on tree phenology in Cameroon and few species have been subjected to detail studies, just as for Tephrosia vogelii and Entada africana. Therefore, the present study was carried out between October, 2012 and September, 2013 to evaluate the phenology of E. africana and T. vogelii in three altitudinal zones in Bambouto Mountains and to understand their response to climatic factors with altitude.
MATERIALS AND METHODS
Study site: This study was conducted on the eastern slope of the Bambouto Mountains (latitude 5°30’ to 5°45’ N, longitude 10°00’ to 10°25’ E and altitude 1400-2500 m) (Fig. 1), Western highlands of Cameroon. This mountain with its maximum height of 2740 m is one of the major volcanic mountains along the complex structure known as the Volcanic Line of Cameroon. Three successive morphological domains separated by more or less pronounced escarpment with distinct environmental conditions were individualized: The upper zone (>2000 m), the mid elevation zone (1600-2000 m) and the lower zone (1400-1600 m)14. The predominant soils of the upper zone are the ando soils while the mid-zone has the andic ferralitic soils. The lower zone has battleship ferralitic soils15. The herbaceous stratum is dominated by Pennisetum purpureum and Imperata cylindrica and ligneous cover is strongly influenced by the anthropogenic activities (agricultural and pasture)16,17.
The climate is the Cameroonian altitude type, with a long rainy season (from March-November) and a short dry season (from December-February). Climatic variables were obtained from the agroclimatic data bases18. Mean monthly rainfall obtain from this data base vary from 8-304 mm with peaks in August and September and the mean monthly temperature vary from 22.2-27°C with peaks in December-March (Fig. 2). The upper zone has a very cool and cloudy climate with temperature ranging between 10-12°C and mean annual rainfall of 2500 mm. The mid elevation zone has a sub-equatorial climate modified by altitude. This climate is cold and humid with an annual average rainfall of 1690 mm and monthly average temperature that is closed to 18°C. The lower zone has warm and humid climate with a monthly average temperature of 23.5°C and annual average rainfall19,20 of 1750 mm.
Experimental procedure: Periodic observations of successive phenophases of the canopy: Leafing, flowering and fruiting were recorded. Trees were marked and were observed with binoculars every 2 weeks during the active growth season and once a month during the dry season from October, 2012 to September, 2013. Phenological observations were made on 120 individuals from two species, 60 individuals, so 20 individuals for each altitude were observed per species.
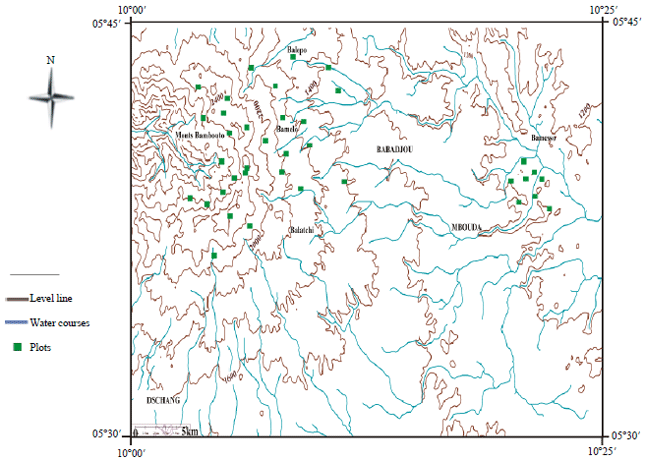 | |
| Fig. 1: | Location of sample trees in the eastern slope of Bambouto mountains |
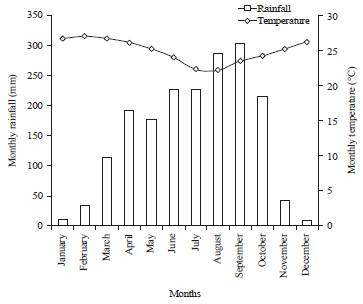 | |
| Fig. 2: | Temperatures and precipitation at the Bambouto mountains |
These species have been selected based on their abundance in the site, their short life cycle, their wide geography distribution and their importance. Ticketing has consisted to affect each individual of each altitude a number (PxY), where Px represent the altitude number and Y represent the individual number. The method use was developed by Grouzis and Sicot6. They proposed for the three phenophases five stages clearly define and easily observation. For foliation the following stages were monitored:
| • | V1: Swelling buds, no leaf development |
| • | V2: Leaf buds and open buds (over 10% and less than 50% of these organs in each individual) |
| • | V3: Leaves mostly open |
| • | V4: Leaves and dry leaves or leaves which have changed colour (over 10% but under 50%) |
| • | V5: Over 50% in each individual with dry leaves and falling leaves |
For flowering the following stages were monitored:
| • | f1: Floral buds only |
| • | f2: Floral buds and open flowers (over 10% and less than 50%) |
| • | f3: Over 50% of organs carrying open flowers |
| • | f4: Open flowers and dry flowers (over 10% and less than 50%) |
| • | f5: A majority of dry flowers and shedding of floral elements |
Fructification was characterized by the following stages:
| • | F1: Early setting stage |
| • | F2: Development of fruit to normal size |
| • | F3: Maturity |
| • | F4: Ripe fruit and onset of dissemination (opening of pods or fall of fruit) |
| • | F5: Fruits dried and fallen |
Stage 1 corresponds to the beginning of the phase and stage 5 to its culmination. Stages 2, 3 and 4 represent a specific phase for each individual, each of the three is characterized by the following degrees of intensity: Low, optimum and declining.
Data analysis: It quantified each of the phenophases for each month of the study as the percentage of plant species that were flowering, fruiting or flushing new leaves. A species was considered in a particular phenophase if one or more of the sampled individuals were in the phenophase. The phenology spectrum is built per calculation for each observation date of population frequencies, individual in phase of foliation (V %), flowering (f %) and fruiting (F %)6. This relation has been used:
![]()
where, P (%) represents the percentage of the individuals of each zone present at diverse phases of foliation (V), flowering (f) or fruiting (F), n represents the number of individuals present at each phases and N is total population. Determinism study of phenological cycles were realized base on phenological data collected, include beginning and ending date of each phenophase (calculated automatically). The inter phenophase duration for the two trees species was calculated.
RESULTS
Leaf-phenology of Entada africana and Tephrosia vogelii: Leaf initiation started at the end of February in E. africana, follows by leaves opening at outset of the month of March at the lower zone, mid elevation zone and at outset of February in the upper zone, continued up to November-December with a peak in May-July (Fig. 3a). Leaf initiation started at the onset of March with return of rainy season in T. vogelii follows by leaves opening at outset of April at the lower zone, mid elevation zone and at the upper zone, continued up to December-January with a peak from May-July (Fig. 4a). This leafing was more precocious for the individuals situated at the upper zone with gap of about 2 weeks in the individuals of the two species in relation to the individuals situated at the low zones.
Leaf-fall: Leaf-fall initiation was a periodic activity in all species, however, the onset of leaf fall initiation varied among the species (Fig. 3a and 4a). Shedding of old leaves was recorded before the start of new ones and the trees were bare for at least a week. In most plant species of E. africana, leaf shedding began in the month of August in the lower zone, mid-zone and mid-August in the upper zone. The T. vogelii shed their leaves in mid-July in the lower zone, mid-zone and in August in the upper zone. The peak in leaf fall was observed from November-December in the two species. Duration of leafing phenophase was long about 12 months in E. africana and 10 months in T. vogelii.
Flowering phenology of Entada africana and Tephrosia vogelii: The intraspecific synchronization was evident for flowering and fruiting with the season rhythm. The flowering activity of E. africana began in the month of March, continued up to the end of June with a peak in May at the three zones (Fig. 3b).
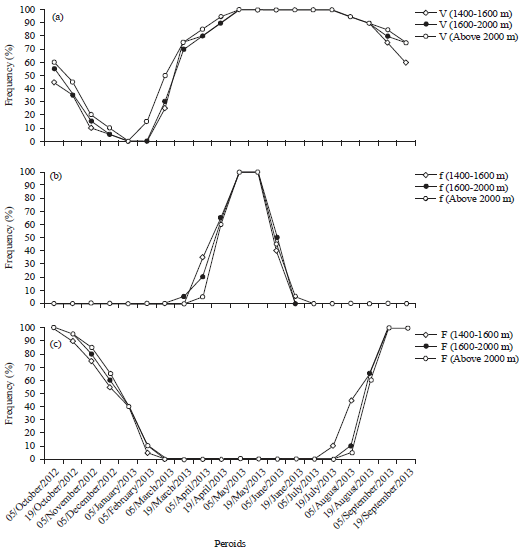 | |
| Fig. 3(a-c): | Phenological periodicity of E. africana in the three altitudinal zones (a) Leafing, (b) Flowering and (c) Fruiting |
The floral buds of T. vogelii had begun to be visible in July in the lower zone, mid elevation zone and in August in the upper zone, continued up to December-January with a peak, respectively in August and September (Fig. 4b). The flowering initiation in two species was much delayed with a gap of 2 weeks for the individuals of E. africana and one month for the individuals of T. vogelii situated above of 2000 m altitude. The duration of the phase was about 3 months in E. africana and 4 months in T. vogelii.
Fruiting phenology of Entada africana and Tephrosia vogelii: Fruiting was followed directly flowering. Young fruits became perceptible with the eyes one month after flowering precisely in the outset of August. The peak period of maturation of fruits was October in E. africana, fruit dehiscence is completed before rainy season (February-March) in all different zones (Fig. 3c). In T. vogelii fruiting activities begins in August at lower zone, mid elevation zone and in September at the upper zone coincide with a peak in October-November. Fruit dehiscence is completed in February (Fig. 4c).
| Table 1: | Approximate inter phenophase duration (in weeks) of two species in Bambouto Mountains |
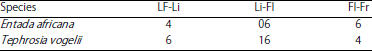 | |
| LF-Li: Leaf fall-leaf initiation, Li-Fl: Leaf initiation-flowering, Fl-Fr: Flowering-fruiting | |
The fruiting in the individuals situated at mid-zone and upper zone was delayed with a gap of 2 weeks at the individuals of the two species in relation to the individuals situated at the lower zone. Duration of fruiting phenophase was long about 8 months in E. africana and 7 months in T. vogelii.
The inter phenophase duration for two woody species is highly variable (Table 1). Leaf-fall and leaf-initiation interphase (leafless period) is long for the two species, indicating the deciduous nature of the study site. The leaf initiation-flowering interphase was also long in T. vogelii excepted E. africana. The E. africana showed long of flowering-fruiting interphase except T. vogelii.
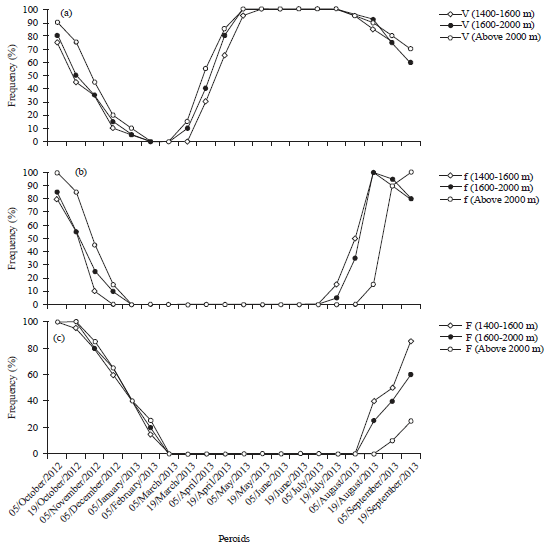 | |
| Fig. 4(a-c): | Phenological periodicity of T. vogelii in the three altitudinal zones (a) Leafing, (b) Flowering and (c) Fruiting |
DISCUSSION
The tropical mountain plants exhibited considerable diversity in leaf initiation, leaf fall, flowering and fruiting activity. However as per climate of the study site, peaks for all phenophases were distinguished due to defined wet and dry seasons. In the three altitudinal zones, leaf initiation peak in May-July, may be attributed to be the hot months of the year before it rains. This may be attributed to the triggering effect of the rising temperature and increase in length of photoperiods7. Rainfall has also significant negative influence indicating that leaves are initiated during the dry season. Leaves production towards the end of the dry season and before the rainy season has also been observed in tropical trees species by Yadav and Yadav7, Kikim and Yadava21 and Singh and Kushwaha22. These authors have suggested that swelling bud before the rainy season would be probably link to a phenomenon of osmotic adjustment due to the liberation of the active osmotic solutes in xylem vessel, they observe ski-lift of bass sap tension in full dry season further to defoliation. Leaf fall is intense during the early dry season because evapotranspiration increases, leading to greater nutrient removal from soil by roots23. The variation in the leaf fall pattern may be due to micro-environmental factors. Similarly, there are reports that leaf fall and bud emergence takes place during the driest part of the year, which coincided with short day length and decrease in temperature and solar radiations24,25. Peak leaf fall during dry season is an important adaptation strategy of the plants of tropical region to minimize the effect of decreased soil moisture or increased water stress conditions7,21,26. The results are also in conformity with Ullmann27, who reported that defoliation observed in dry season goes on well, on the one hand, with pluviometric deficit and mostly with the feedback of minimum level of dry season of the soil water reserve and, on the other hand, with the high temperature could be in favour of leaf fall. Leafing onset by individual E. africana trees in the lower and mid zone was synchronized whereas the individuals in the upper zone showed asynchronous activity. In T. vogelii, leafing onset by individual trees in the mid and upper zone was synchronized whereas the individuals in the lower zone showed asynchronous activity. The precocity observed in the leafing phase apparition in the upper zone in dry season could be attributed to the effects of altitude. Indeed, altitudinal variation involves temperature fluctuation (average of-0.6°C/100 m) and more largely of bio-climate who plays a non-negligible role on the precocity of leafing phase of these species.
A floral bud initiates during the rainy season in the two species have showed that rainfall had positive influence on flowering phenology of these species in the Bambouto Mountains. Rainfall patterns are associated with seasonal cycles of flowering for a number of tropical and subtropical species. In this study, flowering occurred in the transition period between dry and rainy season. This finding is similar to that recorded from other tropical forest in Costa Rican and Mexican tropical dry forest28 and central Borneo29. Others studies have showed a correlation between temperature change and phenological events in tropical plants4,5. Flowering onset by individual of E. africana and T. vogelii trees in Bambouto Mountains was not synchronized, generating a complex flowering population spread over a 3-4 months period. While some tropical species have short and highly synchronized flowering staggered flower initiation may be typical of many tropical tree species with extended flowering periods. Individuals trees within 20 m of each other began flowering on different dates and this difference persisted from year to year. It hypothesized that genotype and trees age are an important factor in determining individual phenological schedules for the two species. Delayed flowering in the upper zone in rainy season in response to the effect of altitude was observed in these species.
Fruiting phenology: Flowering phenophase was related to varying durations of fruiting phenophase (7-9 months). Fruit bud initiates during the rainy season have showed that rainfall had positive influence on fruiting phenology in the Bambouto Mountains. Indeed, Heideman30 and Chapman et al.31 have showed that rainfall influence significantly fruiting, because young fruits in growth require much water for their development. Similarly, there was correlation between fruit production and monthly precipitation. Fruiting phenology in moist forests of northeast Korea corresponds with rainfall periodicity though correlation with rainfall is not explicitly stated21. Tropical evergreen forest of the Western Ghats also showed similar trend with rainfall and fruiting phenology32. Germination during the rainy season is advantageous due to favourable environmental conditions33. It is also the period for the seedling to develop a root system that can withstand the subsequent dry season. Abiotic conditions, seed dispersers, predators and ecological traits of the plant are other factors affecting fruiting and germination phenology5,33. However, the delay observed in apparition of the fruiting in the individuals of the two species situated above of 2000 m altitude could be related to the effects of altitude. It noticed for instance a decrease of temperature of about 23.5-10-12°C while rising from 1400 m to more than 2000 m of altitude20. Synchronization of the phenophases per zone corresponds to the adaptation of these plants in the middle. The asynchronization of the different phenophases between the zones could be imputed to the effects of variations of middle conditions and the altitude.
CONCLUSION
Phenological observations on E. africana and T. vogelii were recorded in terms of different phenophases starting from to fruit dehiscence in three altitudinal zones of Bambouto Mountains. The phenological behaviour of the tree species in this mountain is influenced by rainfall, temperature and altitude. These phenophases of the two species have annual characters. Altitude is responsible to precocity of leafing phase but also the delaying observed in the flowering and fruiting phases of the two species in the upper zone.
SIGNIFICANCE STATEMENT
This study will help the researchers to understanding growth and development of plants in mountains areas in response to rainfall distribution, temperature and altitude are critical to predicting species population responses to climate change.
ACKNOWLEDGMENT
The authors are grateful to the chief of Babadjou village for cooperation and granting permission to do field-work in the Eastern slope of Bambouto Mountains.
REFERENCES
- Haugaasen, T. and C.A. Peres, 2005. Tree phenology in adjacent Amazonian flooded and unflooded forests. Biotropica: J. Biol. Conserv., 37: 620-630.
CrossRefDirect Link - Van Schaik, C.P., J.W. Terborgh and S.J. Wright, 1993. The phenology of tropical forests: Adaptive significance and consequences for primary consumers. Ann. Rev. Ecol. Syst., 24: 353-377.
CrossRefDirect Link - Morellato, L.P.C., D.C. Talora, A. Takahasi, C.C. Becke, E.C. Romera and V.B. Zipparro, 2000. Phenology of Atlantic rain forest trees: A comparative study. BioTropica, 32: 811-823.
CrossRefDirect Link - Marques, M.C., J.J. Roper and A.P.B. Salvalaggio, 2004. Phenological patterns among plant life-forms in a subtropical forest in Southern Brazil. Plant Ecol., 173: 203-213.
CrossRefDirect Link - Grouzis, M. and M. Sicot, 1980. Une methode d'etude phenologique de populations d'especes ligneuses saheliennes: Influence de quelques facteurs ecologiques. Proceedings of the Symposium sur les Fourrages Ligneux en Afrique, April 8-12, 1980, Centre International Pour l'Elevage en Afrique, Addis Abeba, Ethiopia.
- Yadav, R.K. and A.S. Yadav, 2008. Phenology of selected woody species in a tropical dry deciduous forest in Rajasthan, India. Trop. Ecol., 49: 25-34.
Direct Link - Hamann, A., 2004. Flowering and fruiting phenology of a Philippine submontane rain forest: Climatic factors as proximate and ultimate causes. J. Ecol., 92: 24-31.
CrossRefDirect Link - Willis, C.G., B. Ruhfel, R.B. Primack, A.J. Miller-Rushing and C.C. Davis, 2008. Phylogenetic patterns of species loss in Thoreau's woods are driven by climate change. Proc. Natl. Acad. Sci. USA., 105: 17029-17033.
CrossRefDirect Link - Cleland, E.E., J.M. Allen, T.M. Crimmins, J.A. Dunne and S. Pau et al., 2012. Phenological tracking enables positive species responses to climate change. Ecology, 93: 1765-1771.
CrossRefDirect Link - Feteke, F., A. Fayolle, K. Dainou, N. Bourland and A. Die et al., 2017. [Seasonal variations in diameter growth and leaf and reproductive phenology in three Central African commercial timber species]. Bois For. Trop., 330: 3-21, (In French).
CrossRefDirect Link - Morin, S., 1988. Les Dissymetries Fondamentales des Hautes Terres de l'Ouest-Cameroun et leurs Consequences sur l'Occupation Humaine: Exemple des Monts Bambouto. In: L'Homme et la Montagne Tropicale, SEPANRIT (Eds.). Societe pour l'Etude, la Protection et l'Amenagement de la Nature dans les Regions Inter-Tropicales, Talence, France, ISBN: 2-9502879-0-5, pp: 35-56.
- Tematio, P., L. Kengni, D. Bitom, M. Hodson and J.C. Fopoussi et al., 2004. Soils and their distribution on Bambouto volcanic mountain, West Cameroon highland, central Africa. J. Afr. Earth Sci., 39: 447-457.
CrossRefDirect Link - Wouokoue, T.J.B., G.M. Anjah, V.F. Nguetsop and T. Fonkou, 2017. Floristic diversity of the savannah ecosystems in three altitudinal zones of the Bambouto Mountains, West Cameroon. Cameroon J. Biol. Biochem. Sci., 25: 52-59.
Direct Link - Baudoin, W.T.J., N.V. Francois and F. Théophile, 2017. Floristic diversity of Western Highlands Savannas of Cameroon. Int. J. Curr. Res. Biosci. Plant Biol., 4: 7-13.
CrossRefDirect Link - Kikim, A. and P.S. Yadava, 2001. Phenology of tree species in subtropical forests of Manipur in North Eastern India. Trop. Ecol., 42: 269-276.
Direct Link - Singh, K.P. and C.P. Kushwaha, 2005. Paradox of leaf phenology: Shorea robusta is a semi-evergreen species in tropical dry deciduous forests in India. Curr. Sci., 88: 1820-1824.
Direct Link - Morellato, L.P.C., 1992. Sazonalidade e Dinâmica de Ecossistemas Florestais na Serra do Japi. In: Historia Natural da Serra do Japi: Ecologia e Preservacao de Uma Area Florestal no Sudeste do Brasil, Morellato, L.P.C. (Ed.). Universidade Estadual de Campinas/Fundacao de Amparo a Pesquisa do Estado de Sao Paulo, Campinas, Brazil, ISBN-13: 9788526802230, pp: 98-110.
- Borchert, R., 1994. Water status and development of tropical trees during seasonal drought. Trees, 8: 115-125.
CrossRefDirect Link - Borchert, R. and G. Rivera, 2001. Photoperiodic control of seasonal development and dormancy in Tropical stem-succulent trees. Tree Physiol., 21: 213-221.
CrossRefDirect Link - Borchert, R., S.A. Meyer, R.S. Felger and L. Porter-Bolland, 2004. Environmental control of flowering periodicity in Costa Rican and Mexican tropical dry forests. Global Ecol. Biogeogr., 13: 409-425.
CrossRefDirect Link - Brearley, F.Q., J. Proctor, L. Nagy, G. Dalrymple and B.C. Voysey, 2007. Reproductive phenology over a 10-year period in a lowland evergreen rain forest of Central Borneo. J. Ecol., 95: 828-839.
CrossRefDirect Link - Heideman, P.D., 1989. Temporal and spatial variation in the phenology of flowering and fruiting in a tropical rainforest. J. Ecol., 77: 1059-1079.
CrossRefDirect Link - Chapman, C.A., R.W. Wrangham, L.J. Chapman, D.K. Kennard and A.E. Zanne, 1999. Fruit and flower phenology at two sites in Kibale National Park, Uganda. J. Trop. Ecol., 15: 189-211.
Direct Link - Wei, J., 2016. Phenology of tree species in a tropical evergreen forest. Acad. J. Scient. Res., 4: 404-416.
Direct Link - Pao, N.T., K. Upadhaya and A.H. Mir, 2016. Phenological behaviour of tree species in subtropical broad leaved humid forests of Jaintia Hills in Meghalaya, Northeast India. Int. Res. J. Biol. Sci., 5: 10-15.
Direct Link