
Zerihun Jalata
Department of Plant Sciences, College of Agricultural Sciences, Wollega University, Ethiopia
LiveDNA: 251.24518
Firew Mekbib
School Plant Sciences, College of Agriculture and Environmental Sciences, Haramaya University, P.O. Box 138, Dire-Dawa, Ethiopia
LiveDNA: 251.22688
Berhane Lakew
Ethiopian Institute of Agricultural Research (EIAR), Holetta Agricultural Research Center (HARC), P.O. Box 2003 or P.O. Box 20420,
Code 1000, Addis Ababa, Ethiopia
Seid Ahmed
Biodiversity and Integrated Gene Management Program,
International Center for Agricultural Research in the Dry Areas (ICARDA), Morocco
Journal of Applied Sciences
Year: 2018 | Volume: 18 | Issue: 3 | Page No.: 129-137







ABSTRACT
Background and Objective: Scald caused by Rhynchosporium cummune is among the most important disease causing significant yield losses in barley yield. The aim of this study was to investigate the nature of gene actions and combining ability of parents which is useful in designing effective breeding for scald resistant barleys. Materials and Methods: The present study was carried out on 28 barley genotypes (21 F1 crosses and 7 parents) at Holetta, Ethiopia, in two rows of 2.5 m plot length and 0.40 m width at 0.15 m spacing between plants in RCB design with three replications during 2015 main cropping season. Results: The analysis of variance for initial and final percentage ld severity and AUDPC for scald showed highly significant genotypic difference indicating suitability for improvement. The combining ability test showed highly significant variation due to general (GCA) and specific combining ability (SCA) indicating the importance of both additive and non-additive types of gene effects controlling scald resistance. It appears that HB42 and HB1307 with highly significant and negative GCA effects can be promising parents for development of scald resistance and about half of the F1 hybrids also showed negative heterosis estimates over mid parent for all the characters measured. Conclusion: The result suggested that, the resistant additive alleles found in HB42 and HB1307 parents would be fixed using diallel selective or biparental mating of segregants followed by selection.
PDF Abstract XML References Citation
Received: August 04, 2018;
Accepted: October 16, 2018;
Published: November 17, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Zerihun Jalata, Firew Mekbib, Berhane Lakew and Seid Ahmed, 2018. Investigating Parent Combining Ability and Gene Interactions via Diallel Analysis Against Scald Resistance in Barley. Journal of Applied Sciences, 18: 129-137.
DOI: 10.3923/jas.2018.129.137
URL: https://scialert.net/abstract/?doi=jas.2018.129.137
DOI: 10.3923/jas.2018.129.137
URL: https://scialert.net/abstract/?doi=jas.2018.129.137
INTRODUCTION
The fungal pathogen Rhynchosporium commune is anamorphic ascomycete, causing one of the most destructive diseases of barley (Hordeum vulgare L.) especially common in cool temperate climate areas. Infection frequently occurs in the leaf axil, which can lead to chlorosis and eventual death of the leaf1. Barley leaf scald is a polycyclic disease, normally involving several pathogen generations during the growing season and secondary disease spread by splash-dispersed conidia. Scald inflicts yield decreases of up to 40% and reduced grain quality2. Worldwide experiences about scald showed 100% losses in susceptible cultivars under severe epidemics3. In Ethiopia, barley is among important staple crops next to tef, maize, wheat and sorghum mainly grown on about 1 million ha of land with average yield of 2.1 t ha–1 for food, feed and malting4.
However, leaf scald was among widely distributed and destructive diseases in cool highland areas on barley5,6 and yield losses reaching about 67% have been recorded due to scald in Ethiopia7. Hunde et al.6 indicated that the mean grain yield loss assessment due to scald in farmers’ field at Ankober and Faji of central highlands of Ethiopia indicated an average of about 14 and 25% losses in 1999 and 2000, respectively and grain yield losses in malt barley cultivars reach as high as 57% depending on variety and year8. Besides this, a research study reports on various R. cummune isolates collected from different agro-ecologies of Ethiopia indicated divergence in phenotypic and virulence9 and high genetic diversity in the pathogen population and high gene flow between regions and among populations10. This shows barley genotypes that lack broad spectrum resistance to scald may not last long under production. In such case, developing cultivars that combine multiple genes would be a good option to control the scald disease.
Among control measures host plant resistance remains the most dependable and effective means of controlling diseases due to its relatively less costly, biologically safe and convenient for the farmer. Moreover resistance to diseases are important component of integrated disease management programs11. However, the knowledge on the genetic background of the present barley parents was not known which is useful to select suitable parents and effective selection in the development of resistant plants. Many years of research efforts and experiences on screening of barley to scald and net blotch diseases showed limited success12 and hybridization and selection activities were conducted13 which enabled to obtain few resistant cultivars. The damage of scald disease on introduced commercial malting barley cultivars is prevalent. However, the frequent use of resistant few cultivars for long time result an increase of epidemics due to breakdown of resistance. This necessitates resistance breeding continuous activity. The information on the gene actions should be a governing resistance genes and selection of desirable parents for the development of disease resistant varieties is important and this is possible through diallel cross. The combining ability analysis is one of the valuable tools available to ascertain the combining ability effects and helps in selecting the desirable parents14 and it provides the basis for selecting good combiners and also for understanding the nature of gene action15. Study on genetics of scald resistance has been reported. Patil et al.16 indicated that scald resistance alleles were mainly allelic or additive in the NILs (Near isogenic lines) and GCA effects were much stronger than SCA effects. In another report, resistance genes to scald in barley were governed by both major and minor genes that generally have additive effects2 and dominant and recessive genes were also identified17. The study on inheritance of scald resistance on barley lines showed a single dominant gene for resistance18.
In Ethiopia the research reports on barley indicates that scald disease has been among the major biotic stress reducing barley yield and research attempts made so far in breeding for disease resistance to scald was not adequate12,13. Besides this, though most of the exotic germplasms evaluated were highly susceptible to scald but they are used as sources of genes for desirable agronomic traits12. Currently, brewery industries are increasing demanding the necessity of increasing production potential of food or malt barley. Therefore, breeding for disease resistance to scald should be among the research focus of barley improvement to fullfill the gaps in commercial and/or subsistence farming. Hence designing new breeding strategies for gene deployment is a good option to control the yield losses occurring in food and malt barley production and productivity. In this regard, generating information on the nature and magnitude of gene effects controlling inheritance of scald resistance gene and selection of parents would contribute for future resistance breeding.
In view of this, on the basis of their diversity of origin and previous record on the levels of scald resistance, seven barley parents were chosen for this 7×7 half diallel cross of barley. Thus, the aim of this study was to estimate the nature and magnitude of gene actions involved in controlling scald resistance, extent of heterosis and identify desirable parents for future barley scald resistance breeding.
MATERIALS AND METHODS
Experimental site: The study was conducted in 2015 main cropping season at Holetta Agricultural Research Center, which is about 30 km west of Addis Ababa. The area is located at an altitude of 2390 m above sea level (m.a.s.l), latitude of 09°04' N and longitude of 38°30’E (http://www.eiar.gov.et). Holetta is one of hot spot area for scald and net blotch barley disease where most of screening has been done in Ethiopia.
Planting materials and experimental design: Seven barley cultivars (Table 1) with different levels of resistance and susceptibility to scald disease and origin were used in half diallel crossing in 2014/15 main cropping season to generate 21 F1 crosses for field evaluation. The F1 crosses were obtained by hand emasculation and pollination in the field. Then a total of 28 genotypes including seven parents and 21 F1 crosses were planted at Holetta in a randomized complete block design with three replications during the 2015 main cropping season. Seeds of each genotype were sown in two rows of 2.5 m length and 0.40 m width at 15 cm between plants. To increase the disease epidemics, the spreader rows of scald susceptible variety, Sabini, was planted surrounding each block and plot.
Scald assessment: Scald disease severity was scored on ten randomly selected plants in each plot using double digit scale (D1D2, 00-99) based on Saari and Prescott19 in the field under natural condition. The first digit (D1) indicates vertical disease progress on the plant and while the second digit (D2) refers to severity measured as diseased leaf area. Disease scoring was repeated weekly since 10, September, 2015 for five times starting at 53 growth stages20 as modified by Tottman and Makepeace21. The plot mean severity scores of each plot was converted into percentage severity scale for all the growth stages such that for each score, the percentage of disease severity was estimated using the formula of disease severity22:
![]()
Area under disease pressure curve (AUDPC) was calculated to estimate the scald severity over time based on the five growth stages’ percent disease severity estimations according to Shaner and Finney23 formula. It was computed as:
![]()
where, Yi is the disease severity on the ith date, Ti+1-Ti is time or days between two disease scores, n = number of dates on which the disease was recorded.
Data analysis: One way analysis of variance (ANOVA) with significance level 0.01 and 0.05 was performed using the general linear model (GLM) procedure in the SAS version 9.1 software24. Significance test of the characters were determined before diallel analysis. Then diallel analysis were conducted according to Griffing25 method 2 and model 1(fixed effect) using the SAS program for Griffing’s diallel analysis by Diallel SAS program of Zhang and Kang26 and Zhang et al.27. Mid parent heterosis (MPH) and better parents heterosis (BPH) or heterobeltiosis estimated as outlined by Falconer and Mackay28 and significance test was done according to Wynne et al.29.
RESULTS
The analysis of variance for initial percent severity, final percent severity and AUDPC for scald showed highly significant (p<0.01) genotypic differences (Table 2). The mean performance estimates for the barley genotypes varied for initial percent severity, final percent severity and AUDPC of scald were indicated in Table 3, 4 and Fig. 1. Sabini×HB42, Grace×HB42, Misrach×HB42, Miscal-21×HB42, HB1307×HB42, HB42 and HB42×Agegnehu genotypes showed relatively the lowest scores for initial percentage severity, final percentage of severity and AUDPC for scald (reduced symptoms and increasing resistance).
| Table 1: | Description of seven barley cultivars used in half diallel crossing in 2014 |
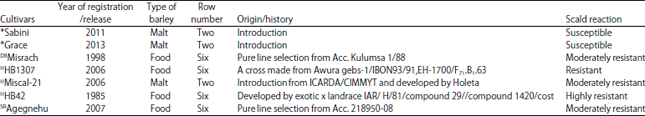 | |
*Introduced (personal communication with Dr. Berhane Lakew), DBDeveloped by Holetta Agricultural Research Center and released by Debre Berhan Agricultural Research Center, HReleased and developed by Holetta Agricultural Research Center, SRReleased and developed by Sirinka Agricultural Research Center. Sources30,31 | |
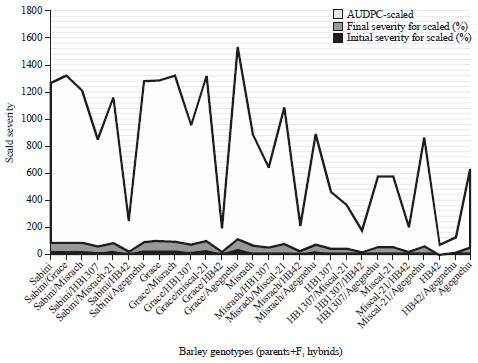 | |
| Fig. 1: | Mean performances of 28 barleys genotypes for initial percentage scald severity, final percentage scald severity and AUDPC for scald disease |
| Table 2: | Analyses of variance for percent initial scald severity, percent final severity and AUDPC for scald in 2014/15 cropping season |
 | |
| *,**Significance at the 0.05 and 0.01 probability level, respectively, GCA: General combining ability, SCA: Specific combining ability, h2: Heritability, CV: Coefficient of variation | |
On the contrary, Sabini and Grace were the most susceptible parents and Fl hybrids obtained from crossing of these parents, Sabini and Grace showed higher AUDPC scald scores except with HB42 parent. The correlation between the initial percentage scald severity with final percentage severity (r = 0.764**) and with AUDPC (r = 0.892**) was highly significant, the association between final scald severity and AUDPC for scald was also high (r = 0.950**).
The analysis of variance for combining ability showed highly significant (p<0.01) general combining ability (GCA) and specific combining ability (SCA) for initial percentage scald severity, final percentage scald severity and AUDPC as percentage days (Table 2). The Baker ratio and narrow sense heritability was closer to unity for percentage initial scald severity, percentage final scald severity and AUDPC for scald. The GCA effects for initial percentage scald severity, final percentage scald severity and AUDPC for scald is displayed in Table 3. In the present study, HB42 and HB1307 parents showed highly significant negative GCA effects for initial percentage severity, final percentage severity and AUDPC for scald. While Miscal-21 with significant GCA effect for initial percentage severity also showed negative AUDPC for scald. SCA effects, mid parent heterosis (MPH) and better parent heterosis (BPH) for percent initial scald severity, percent final scald severity and AUDPC as percent days is indicated in Table 4.
| Table 3: | Estimated general combining ability (GCA) effects of parents and their corresponding mean performance for initial percentage scald severity, final percentage scald severity and AUDPC as percentage days for scald disease in 2014/15 cropping season |
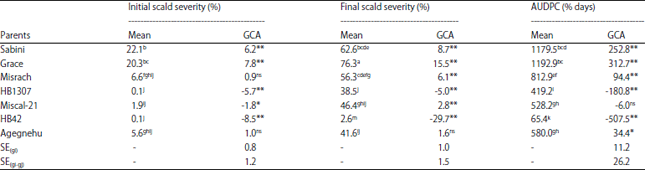 | |
*,**Significance at the 0.05 and 0.01 probability level, respectively, ns: Non significant, means with the same letter are not significantly different at alpha 0.05 level of significance, CV: Coefficient of variation, SE(gi): Standard error of GCA, SE(gi-gi): Standard error of GCA for two GCA effects | |
From 21 crosses, nearly half of the crosses showed negative SCA effects for initial percentage scald severity, final percentage scald severity and AUDPC as percent for scald.
The F1 barley hybrids showed negative and highly significant heterosis for scald severity (11 crosses), final percent scald severity (8 crosses) and AUDPC as percent days (9 crosses) over mid parent. Whereas negative heterosis was observed on five and one barley hybrid for initial percentage scald severity and AUDPC as percent days for heterosis over better parent, respectively (Table 4).
DISCUSSION
In resistance breeding identifying sources of resistance and the effective breeding systems to use is an important consideration to design an effective breeding. In this study though a one year data may inflate some of the results but the experiment was conducted in hot spot area for the disease and the experimental parents may provide preliminary information on the exploitation of the resistance sources in the future barley breeding. Thus the analysis of variance for initial percentage severity, final percentage severity and AUDPC for scald showed highly significant (p<0.01) genotypic differences (Table 2) indicating the presence of broad genetic potential for selecting scald resistant plants. For most of barley genotypes, final severity symptom and AUDPC for scald were associated with the amount of initial percentage severity which showed high initial scald severity produced high final percentage severity and AUDPC for scald. AUDPC value can be calculated from two data points but greater accuracy is achieved with each additional observation32. With multiple severity readings, the AUDPC has been calculated as a measure of slow rusting resistance33. The AUDPC has been widely used to measure whether rusting was fast or slow while vertical resistance in the host is effective against initial inoculums or delays the start of an epidemics34 and quantitative resistance shows no obvious genetic interaction with the pathogen and slows down the disease development by increasing latency period and other parameters related to the epidemic35. It was observed that there was a difference among barley genotypes in reducing the initial and/or final scald disease epidemics which may be used as a possible indicator of the type of resistance the genotype contains.
The highly significance (p<0.01) of GCA and SCA for initial percentage scald severity, final percentage scald severity and AUDPC as percent days (Table 2) for parents and crosses revealed that there was high involvement of additive gene actions (fixable) and non-additive gene actions controlling scald resistance. Baker36 suggested that the progeny performances could be predicted by the use of the combining ability ratio. In this study, the Baker ratio and narrow sense heritability was closer to unity for all parameters measured (percentage initial scald severity, percentage final scald severity and AUDPC for scald) (Table 2) confirmed the predominance of additive gene effects to non-additive gene effects in controlling the inheritance of scald resistance. Moreover, the broad sense heritability estimates for the same traits were relatively high (Table 2) implying high possibility of success in fixing scald resistance genes through selection during early generation. The additive variance is the main determinant of the observable genetic properties of the population and selection response28. High genetic advances could be realized when working on traits with higher additive genetic variance36,37. In other studies on inheritance of scald resistance, the development of near isogenic lines (NILs), BC7 generation of barley with the recurrent parent, scald resistance alleles were mainly allelic or additive in the NILs and GCA effects was much stronger than SCA effects16.
| Table 4: | SCA effects, mid parent heterosis (MPH) and better parent heterosis (BPH) for percentage initial scald severity, percentage final scald severity and AUDPC as percentage days on barley hybrids in 2014/15 cropping season |
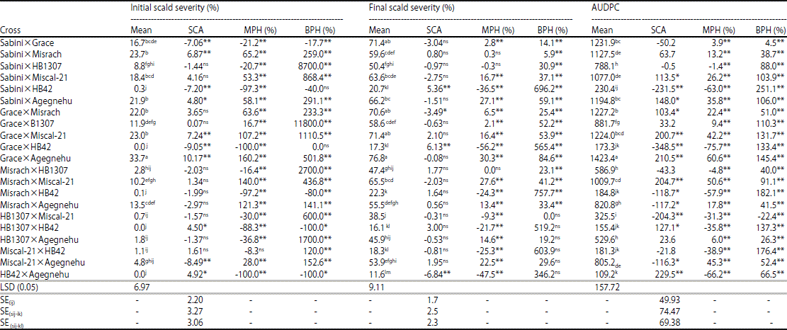 | |
| *,**Significance at the 0.05 and 0.01 probability level, respectively, ns: Non significant, SE(Sij): Standard error for testing all SCA, SE(Sij-ik): Standard error for comparing the difference between two SCA having one common parent, SE (Sij-kl): Standard error for testing the SCA of two crosses having no parent in common, means in the same column followed by the same letter are not significantly different | |
Similarly resistance genes to scald in barley were governed by both major and smaller minor genes that generally have additive effects2. In another research dominant gene and recessive genes were reported17. This result is partly coincides with Singh et al.18 report which indicated the inheritance of scald resistance in segregating barley generations showed a single dominant gene. Similarly, broad sense heritability of 0.64 of scald resistance to all scald isolates was reported38 and high broad sense heritability of 0.850 and 0.967 was obtained in two barley crosses39. Heritability in broad sense estimates the genetic proportion (additive dominance and interaction component) of the total phenotypic variation, while heritability in narrow sense estimates only the additive portion. Its high estimates show a predominance of additive genetic variance in their expression40.
Combining ability provides the basis for selecting good combiners and also for understanding the nature of gene action15. A parent with a significant negative GCA value would contribute a high level of disease resistance and whereas a parent with a positive value would contribute a high level of susceptibility41. Thus, both HB42 and HB1307 parents contributed in transferring scald resistance genes to their progenies. Furthermore, comparison between HB42 and HB1307 with GCA effects in the same direction showed that GCA effects of both HB42 and HB1307 parents were highly significantly different from each other at 1% level suggesting that HB42 was the best and superior general combiner for increasing scald resistance. According to Griffing25 high GCA effects are mostly due to additive gene effects or additive x additive interaction effects. In this regard, these two parents with good general combiners can be useful in breeding for scald resistance.
It is believed that a cross between two good combiners results in a hybrid with high SCA. On the contrary, a cross between poor parents may also result in a better hybrid or in other words may have significant SCA42. In this study, the combinations between poor×best (Grace×HB42/Sabini× HB42, best×poor combiners (HB1307×Miscal-21) and best× best (HB42×HB1307) combiners for initial percent scald severity, final percent scald severity and AUDPC as percent days resulted in hybrids with either significantly negative SCA effects and/or negative MPH or BPH. A hybrid combination between best x poor combiners (HB1307×Miscal-21) was the only F1 hybrid that showed negative heterosis over better parent for AUDPC for scald even superior to a hybrid combination of the two good parents (HB1307×HB42). Though a hybrid HB1307×HB42 had positive SCA effects for all the characters studied, it exhibited highly significant and negative MPH for the characters measured which is contradictory. In general it appears that there is better chance of producing better resistant progeny from combination of resistant parents.
Commercial cultivars and/or barley lines introduced from abroad for research or commercial purpose were usually observed susceptible to diseases and less adapted in Ethiopia. Recently introduced malt barley cultivars such as Sabini and Grace used in this research are examples of such phenomenon. The absence of adaptable high yielding commercial malting barley cultivars in Ethiopian barley landrace has been limited. As a result, an introduction of commercial cultivars or germplasm from abroad is a must to fill the gap. Thus, this study tried to shed light on the possibility of developing resistant, adaptable cultivars and/or lines with high yielding potential using the Ethiopian barleys such as HB42 and HB1307 as sources of resistance genes to scald. Hence to bring back the susceptible cultivars of high yielding potential to production, it is suggested that transferring resistance genes from either HB42 and/or HB1307 to Sabini and Grace malt barley cultivars which are important but susceptible to scald.
CONCLUSION
The combining ability analysis showed highly significant GCA and SCA among barley genotypes tested for initial, final percent severity and AUDPC as percentage of days indicating the involvement of additive and non-additive types of gene actions governing the inheritance of scald resistance. Both HB42 and HB1307 parents were best general combiners for scald resistance. Furthermore, nearly half of the total crosses showed negative heterosis for scald over mid parent. The result suggested that the resistant additive alleles would be fixed using diallel selective or biparental mating of segregants followed by selection.
ACKNOWLEDGMENT
The authors would like to thank the Ethiopian Ministry of Education, Haramaya University and Wollega University for their financial support to conduct this research. The Ethiopian Institute of Agricultural Research, Holeta Agricultural Research Center, Barley Breeding Program, is also acknowledged for providing research facilities, experimental land and cooperation to carry out all research activities and field evaluation.
REFERENCES
- Avrova, A. and W. Knogge, 2012. Rhynchosporium commune: A persistent threat to barley cultivation. Mol. Plant Pathol., 13: 986-997.
CrossRefDirect Link - Zhan, J., B.D.L. Fitt, H.O. Pinnschmidt, S.J.P. Oxley and A.C. Newton, 2008. Resistance, epidemiology and sustainable management of Rhynchosporium secalis populations on barley. Plant Pathol., 57: 1-14.
CrossRefDirect Link - Yitbarek, S., L. Berhane, A. Fikadu, J.A.G. van Leur, S. Grando and S. Ceccarelli, 1998. Variation in Ethiopian barley landrace populations for resistance to barley leaf scald and netblotch. Plant Breeding, 117: 419-423.
CrossRefDirect Link - Alemu, T.W., B. Hundie and W.T. Bayisa, 2015. Evaluation of integrated disease management for malt barley production in bale highlands, South-Eastern Ethiopia. Global J. Pests Dis. Crop Protect., 3: 108-112.
Direct Link - Meles, K., M. Hulluka and M.M. Abang, 2004. Phenotypic diversity in Rhynchosporium secalis from Ethiopia and host response to barley scald. Plant Pathol. J., 3: 26-34.
CrossRefDirect Link - Selvaraj, C.I., P. Nagarajan, K. Thiyagarajan, M. Bharathi and R. Rabindran, 2011. Studies on heterosis and combining ability of well known blast resistant rice genotypes with high yielding varieties of rice (Oryza sativa L.). Int. J. Plant Breed. Genet., 5: 111-129.
CrossRefDirect Link - Rajendran, A., A. Muthiah, J. Joel, P. Shanmugasundaram and D. Raju, 2014. Heterotic grouping and patterning of quality protein maize inbreds based on genetic and molecular marker studies. Turk. J. Biol., 38: 10-20.
Direct Link - Patil, V., A. Bjornstad, H. Magnus and J. Mac Key, 2002. Resistance to scald (Rhynchosporium secalis) in barley (Hordeum vulgare L.). II. Diallel analysis of near‐isogenic lines. Hereditas, 137: 186-197.
CrossRefDirect Link - Singh, A.K., B.G. Rossnagel, G.J. Scoles and R.A. Pickering, 2003. Inheritance of scald resistance from barley lines 4176/10/n/3/2/6 and 145L2. Can. J. Plant Sci., 83: 417-422.
CrossRefDirect Link - Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for the growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link - Tottman, D.R. and R.J. Makepeace, 1979. An explanation of the decimal code for the growth stages of cereals, with illustrations. Ann. Applied Biol., 93: 221-234.
Direct Link - Sharma, R.C. and E. Duveiller, 2007. Advancement toward new spot blotch resistant wheats in South Asia. Crop Sci., 47: 961-968.
Direct Link - Shaner, G. and R.E. Finney, 1977. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in knox wheat. Phytopathology, 67: 1051-1056.
CrossRefDirect Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Zhang, Y., M.S. Kang and K.R. Lamkey, 2005. DIALLEL-SAS05: A comprehensive program for Griffing's and Gardner-Eberhart analysis. Agron. J., 97: 1097-1106.
CrossRefDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Wynne, J.C., D.A. Emery and P.W. Rice, 1970. Combining ability estimates in Arachis hypogaea L. II. Field performance of F1 hybrids. Crop Sci., 10: 713-715.
CrossRef - Abtew, W.G., B. Lakew, B.I. Haussmann and K.J. Schmid, 2015. Ethiopian barley landraces show higher yield stability and comparable yield to improved varieties in multi-environment field trials. J. Plant Breed. Crop Sci., 7: 275-291.
Direct Link - Jeger, M.J. and S.L.H. Viljanen-Rollinson, 2001. The use of the area under the disease-progress curve (AUDPC) to assess quantitative disease resistance in crop cultivars. Theor. Applied Genet., 102: 32-40.
CrossRefDirect Link - Cisar, G., C.M. Brown and H. Jedlinski, 1982. Diallel analyses for tolerance in winter wheat to the barley yellow dwarf virus. Crop Sci., 22: 328-333.
CrossRefDirect Link - Feriani, W., M. Cherif and S. Rezgui, 2012. Preliminary genetic analysis of resistance to scald in Tunisian barley. Int. J. Plant Breed., 6: 95-99.
Direct Link - Aoki, E., T. Baba, O. Yamaguchi, S. Ito and J. Moriwaki, 2011. Development of barley cultivars with resistance to scald (Rhynchosporium secalis (Oud.) Davis) in Japan. Japan Agric. Res. Quarterly, 45: 349-357.
CrossRefDirect Link - Chakraborty, S., H.K. Borah, B.K. Borah, D. Pathak, B.K. Baruah, H. Kalita and B. Barman, 2010. Genetic parameters and combining ability effects of parents for seed yield and other quantitative traits in black gram [Vigna mungo (L.) Hepper]. Notulae Sci. Biol., 2: 121-126.
CrossRefDirect Link - Hakizimana, F., A.M. Ibrahim, M.A. Langham, S.D. Haley and J.C. Rudd, 2004. Diallel analysis of wheat streak mosaic virus resistance in winter wheat. Crop Sci., 44: 89-92.
CrossRefDirect Link