
Eman A. ABO Elfadl
Department of Animal Husbandry and Wealth Development, Faculty of Veterinary Medicine, Mansoura University, El-Gomhouria Street, 35516 Mansoura, Dakahlia, Egypt
Hend A. Radwan
Department of Animal Husbandry and Wealth Development, Faculty of Veterinary Medicine, Mansoura University, El-Gomhouria Street, 35516 Mansoura, Dakahlia, Egypt
Journal of Applied Sciences
Year: 2016 | Volume: 16 | Issue: 8 | Page No.: 380-387







ABSTRACT
Objective: The study aimed at selecting the accurate model helps in improving productive traits in dairy population through compare between inclusion and exclusion of maternal genetic effect in estimating variance components and genetic parameters of those traits. Methodology: Data set and pedigree information were collected from 3464 Holstein-Friesian dairy records for 991 cows descended from 99 bulls and 691 dams calved over a period of 17 years (1994-2011) for three consecutive lactations. Three multi-trait animal models were fitted to estimate covariance components and corresponding genetic parameters. Results: The highest direct and maternal genetic variances were in model 2 which account for covariance between direct and maternal genetic effect. Mostly, phenotypic variances were high in full animal model for first and second lactations but in third lactation the highest being in model 3. Direct and maternal heritability were high in model 2 in three lactations except direct heritability for days in milk in second lactation and days in milk, 305-day milk yeild mature equivalent, fat yield and protein yield in third lactation. Ranges and standard deviations of breeding value varied between different models but often they were high in model 1 and 3. Conclusion: Results highlight the importance of inclusion of maternal genetic effect in designing an appropriate selective and breeding program to obtain accurate estimates of genetic parameters especially if the variation between animals is high.
PDF Abstract XML References Citation
Received: June 07, 2016;
Accepted: June 22, 2016;
Published: February 25, 2019
How to cite this article
Eman A. ABO Elfadl and Hend A. Radwan, 2016. Model Comparisons and Genetic Parameters Estimates of
Productive Traits in Holstein Cows. Journal of Applied Sciences, 16: 380-387.
DOI: 10.3923/jas.2016.380.387
URL: https://scialert.net/abstract/?doi=jas.2016.380.387
DOI: 10.3923/jas.2016.380.387
URL: https://scialert.net/abstract/?doi=jas.2016.380.387
INTRODUCTION
Dairy calves are usually separated from their mothers soon after birth. Thereby any maternal effect can be assumed to have occurred prenatally1-3. Prenatal effects include maternal additive genetic effect which mean inherent ability of the mother to provide a suitable environment to embryo plus maternal permanent environment effect which include both dam’s mothering ability and the maternal non additive genetic effect of the dam3.
Maternal effects have been defined as any influence from a dam on its offspring, excluding the effects of directly transmitted genes that affect performance of the offspring4. In mammals, environmental variation in the offspring is partially due to genetic variation of some other traits from the dams, such that quantitative traits can be influenced by two genetic components, animal genotype (direct genetic effect) and dam genotype (maternal genetic effect)5. In recent years, some published reports demonstrated that one of the maternal effects such as additive maternal genetic and permanent environmental effects (or both) could significantly affect milk yield traits.
Many factors affect milk, fat and protein yield including direct genetic effect, maternal genetic effect and environmental effects. Assessment of phenotypic and genetic variations between individuals has a great role in the determination of direct additive genetic value of milk production traits in cattle population as reviewed by Pantelic et al.6.
Both direct and maternal effects should be taken into consideration to realize the optimum genetic improvement in selection programs as showed by Robison7. Success in designing breeding strategies can be evaluated through actual change in additive genetic value (breeding value) as the proportion of predicted change of the breeding value mean for the same trait under condition as mentioned by Jurado et al.8.
Inheritance of economic traits in dairy cows is cytoplasmic rather than nuclear in origin, so heritability from daughter dam regressions are higher than those estimates from paternal half-sib analysis9,10. Cytoplasmic origin was a significant source of variation in yield traits of dairy cattle11. Huizinga et al.12 attributed 13% of variation in milk economic returns and 10% of variation in milk, fat and protein yields to cytoplasmic components.
Genetic evaluations of dairy cows for some economic traits have been shown to depend mainly on estimation of variance, covariance, heritability, genetic and phenotypic correlations essential for predicting correlated and direct response to selection as reported by Van Vleck13.
This study was conducted to compare three models in estimating genetic parameters, variance and covariance components for some productive traits in first three lactations in Holstein-Friesian cattle based on inclusion and/or exclusion of direct maternal effect through the variance component estimation restricted maximum likelihood (VCE- REML).
MATERIALS AND METHODS
This study was carried out through a field survey of 3464 dairy records for 991 Holstein-Friesian cows, daughters of 691 dams and 99 sires belonging to a commercial dairy herd (Alexandria-Copenhagen Company), situated in Cairo-Alexandria desert road and about 76 km far from Alexandria, over a period of 17 years (1994-2011). All animals were kept in an open system, supplied with a cool spraying system during hot climate. Animals had a free access to clean water. The cows were fed on Total Mixed Ration (TMR) according to their production state. The ingredient composition of rations were formulated according to National Research Council (NRC, 1916) and the TMR composition was 30% corn silage, 28.6% alfalfa, 14.7% corn, 18-19% crude protein, 7% glutein, 6.4% linseed meal, 4.6% soya beans meal, 3% cotton seed meal, 1.3% wheat bran, 1.2% hay, 0.4% calcium carbonate, 0.3% sodium bicarbonate and 0.3% sodium chloride.
All cows were machine milked 3 times a day with 8 h interval between milkings. Cows were dried-off about 60 days before expected calving date or abruptly at 210 days of pregnancy. Investigated traits were days open of cows per day (DO), days in milk of cows per day (DIM), total milk yield per kg (TMY), 305-day milk yield per kg (305-day MY), fat yield per kg (FY) and protein yield per kg (PY). Data were analyzed statistically, variance components and genetic correlation among traits were estimated via REML applying the package VCE according to Groeneveld et al.14. Direct and maternal heritability estimates were calculated using multi-trait analysis. Three models were used to estimate covariance components and corresponding genetic parameters of the studied traits. Tested models were the follows:
| • | Model 1: Y = Xb+Z1a+e |
| • | Model 2: Y = Xb+Z1a+Z2m+e with Cov (am)#0 |
| • | Model 3: Y = Xb+Z1a+Z2m+e with Cov (am) = 0 |
where, Y is a vector of records for studied traits; b, a, m and e are vectors of fixed, direct genetic, maternal genetic effects and the residual effects, respectively. The X, Z1 and Z2 are corresponding design matrices associating the fixed, direct genetic, maternal genetic effects to vector of Y. With assumption of direct additive genetic, maternal additive genetic and residual effects are normally.
The model had the following distributional assumptions:
E (Y) = Xb, E (a) = 0, E (m) = 0, E (e) = 0
The inverse of the numerator relationship matrix (A–1) was considered and the variances and co variances were:
Var (a) = A σ2a, Var (m) = A σ2m, Var (e) = I σ2e
where, σ2a, σ2m and σ2e are variances due to direct additive genetic, maternal genetic effect and random error, respectively. And A is the additive genetic relationship matrix and I is the identity matrix15. Standard errors of genetic correlations were calculated using the approximate formula according to Robertson16 and Falconer and Mackay17.
RESULTS
A summary of the data and descriptive statistics of all traits in the data set used for parameters estimation in the three lactations are presented in Table 1. The means of yield traits ranged from 8625, 8098, 264.1 and 219 kg for TMY, 305-day MY, FY and PY, respectively to 8950, 8801, 268 and 220 kg, respectively. Coefficient of variation (CV%) were in the range of 35.48, 25.69, 37.61 and 38.03%, respectively to 40.25, 31.19, 42.63 and 41.67%.
Variance covariance components and heritability estimates for all traits in three lactations are given in Table 2-4, respectively. Additive genetic variances were higher in model 2 than those for both model 1 and model 3 for the studied traits in all lactations except for DIM, TMY, 305-day MY, FY and PY in the second lactation. Maternal genetic variances were higher in model 2 than those for both model 1 and model 3 for the studied traits in all lactations. However, residual and phenotypic variances were higher mostly in model 1 and model 3 than their corresponding values in model 2.
| Table 1: | Least squares means, standard deviations and coefficients of variation for different productive traits in three lactations of Holstein cows |
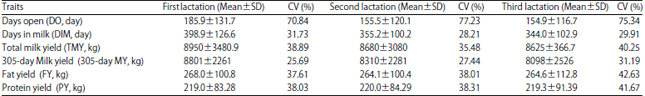 | |
| SD: Standard deviation, CV%: Coefficient of variation per percent | |
| Table 2: | Variance components of the first lactation order in Holstein cows |
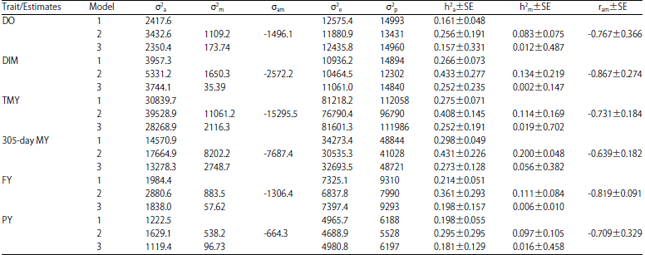 | |
DO: Days open, DIM: Days in milk, TMY: Total milk yield, 305-day MY: 305-day milk yield, FY: Fat yield, PY: Protein yield, σ2a: Direct additive genetic variance, σ2m: Maternal additive genetic variance, σam: Direct maternal genetic covariance, σ2e: Residual variance, σ2p: Phenotypic variance, h2a: Direct heritability, SE: Standard error, h2m: Maternal heritability and ram: Direct maternal genetic correlation | |
| Table 3: | Variance components of the second lactation order in Holstein cows |
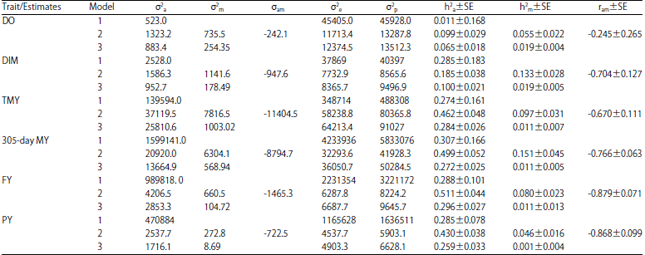 | |
DO: Days open, DIM: Days in milk, TMY: Total milk yield, 305-day MY: 305-day milk yield, FY: Fat yield, PY: Protein yield, σ2a: Direct additive genetic variance, σ2m: Maternal additive genetic variance, σam: Direct maternal genetic covariance, σ2e: Residual variance, σ2p: Phenotypic variance, h2a: Direct heritability, SE: Standard error, h2m: Maternal heritability and ram: Direct maternal genetic correlation | |
| Table 4: | Variance components of the third lactation order in Holstein cows |
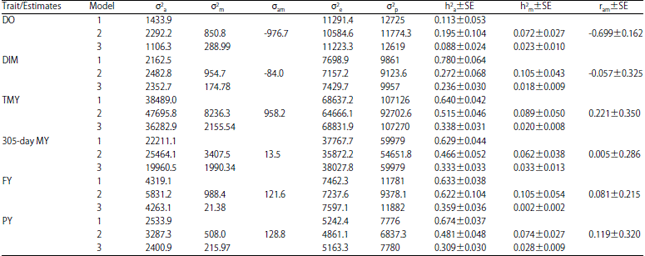 | |
DO: Days open, DIM: Days in milk, TMY: Total milk yield, 305-day MY: 305-day milk yield, FY: Fat yield, PY: Protein yield, σ2a: Direct additive genetic variance, σ2m: Maternal additive genetic variance, σam: Direct maternal genetic covariance, σ2e: Residual variance, σ2p: Phenotypic variance, h2a: Direct heritability, SE: Standard error, h2m: Maternal heritability and ram: Direct maternal genetic correlation | |
Regarding heritability, direct and maternal heritabilities were higher in model 2 than those obtained from model 1 and 3 for all studied traits in all lactations except DIM, TMY, 305-day MY, FY and PY in the third lactation and DIM in the second lactation for direct heritability. Direct maternal genetic covariances (σam) and correlations (ram) were negative for the studied traits in all lactations except TMY, 305-day MY, FY and PY in the third lactation.
Estimates of breeding values for the traits under study in all lactations are given in Table 5. High ranges of estimated breeding values were mostly obtained from model 1 and 3 for the studied traits in all lactations except DO and DIM those were higher in model 2 in the third lactation. Spearman correlation coefficients of breeding value among different models for the studied traits in three lactations are presented in Table 6. Positive and negative coefficients were significant among different models except model 1 and 2 for TMY, 305-day MY, FY and PY in the second lactation as well as model 1 and 3 for DIM, FY and PY in the second lactation were non-significant.
| Table 5: | Maximum, minimum, standard errors and range of estimated breeding values of milk productive traits for all animals obtained from multi statistical animal model analysis for first, second and third lactation in Holstein cows |
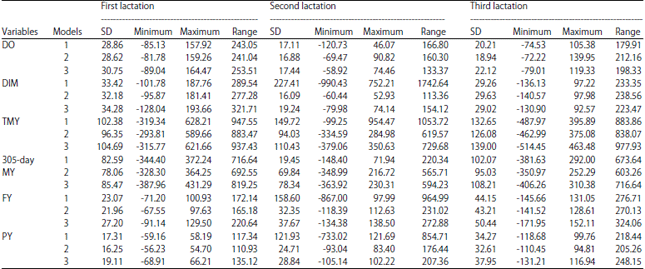 | |
| DO: Days open, DIM: Days in milk, TMY: Total milk yield, 305-day MY: 305-day milk yield, FY: Fat yield, PY: Protein yield and SD: Standard deviation | |
| Table 6: | Spearman correlation coefficients of the first, second and third lactation in Holstein cows |
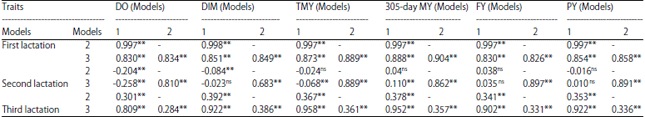 | |
| DO: Days open, DIM: Days in milk, TMY: Total milk yield, 305-day MY: 305-day milk yield, FY: Fat yield, PY: Protein yield, ns: Non-significant, **p<0.01 | |
DISCUSSION
The primary objective of this study was to compare among different models to show the importance of inclusion or exclusion of maternal genetic effect during formulation selecting and breeding planes for future genetic improvement.
In this study the overall adjusted means for days open were higher than those reported by Kadarmideen et al.18 being 107 days but lower than 162.7 days obtained by Shalaby19 compared to second and third lactations in the present study. Days open is considered an indicator for reproductive performance which has high unfavorable values in this study indicating that high producing cows might have been exposed to negative energy balance at re-breeding time.
Regarding days in milk, the overall adjusted means for DIM were higher than those calculated by Shalaby et al.20 being 319.8±128 days in early lactation in Friesian cattle in Egypt and Shalaby19 being 353.9±93.5 days in commercial herds in Egypt compared to second and third lactations of the present study. The CV% for DIM in three lactations were higher than those reported by Alemam21 being 24.7%. High CV% could indicate possibility for a genetic improvement.
The current means and standard errors for TMY in three lactations were lower than those obtained by Hussain et al.22 being 1194±9, 1311±10 and 1354±10 kg, respectively. High milk yield usually indicates good caring and nutrition programs.
Kadarmideen et al.23 calculated means and SD for 305-day MY, fat yield and protein yield to be lower than those calculated in the present study for 305-day MY (6851±1599 kg) and higher fat (270±65 kg) and protein (224±52 kg) yield, respectively in UK dairy cows. Furthermore, Konig et al.24 calculated means and SD for 305-day MY and DO to be 3867±1257.6 kg and 129.5±64.3 days on 2764 and 1623 in Northern Thai dairy cattle, respectively.
In the same line with the present study, Schutz et al.11 applied different animal models on milk production traits in dairy cattle to compare among full animal model, full animal model with covariance and without covariance between direct and maternal genetic effect. Additive genetic variances for 305-day MY were 575,951, 323,483 and 583,040 kg for previous models, respectively. While, additive variance for FY were 1130, 1179 and 1130 kg, respectively. The previous results of Schutz et al.11 contradicted the results of the present study in which the highest variance components were detected in model 2 for first and third lactations and in model 1 for second lactation. Most of productive traits not only depend on genetic potential of an animal but also on maternal effect plus environment either permanent or temporary.
Similarly, maternal genetic variances for 305-day MY were 59,609 and 14 kg for models accounted for cov (am) = Aσam and model account for cov (am) = 0, respectively. For previous productive traits maternal effects are essential in case of selection for these traits especially if the covariance between animals is high. While, maternal genetic variance for fat yield was 1 in both models (model 2 and 3), but here in this study the highest was obtained in model 2.
Phenotypic variances for 305-day MY and FY were contradicted the present study, the highest yields were 2,314,464 and 3448 kg found in model with cov (am)#0.0, respectively. In the present study, if maternal effect was present but not considered (cov (am) = 0), phenotypic variances could have been higher than if maternal effects were considered but both of them were lower than additive direct and phenotypic variances which contain at least part of maternal effect. This finding is agree to that of Meyer25 who reported that models that do not depend on maternal genetic effect would lead to an increase in estimates of additive genetic variance and direct heritability.
Furthermore, Schutz et al.11 estimated residual variance for FY to be in agreement with that of the present study in first and third lactations, the highest fat yield was 1299 kg obtained in model with cov (am) = 0.0. Indicating that maternal effect should be considered during formulation of selecting plans even if there is no variation among animals. However, residual variance for 305-day MY was different from this study that the highest variance being 981,646 kg obtained in model with cov (am)#0. Maternal effect is essential during designing an efficient breeding program therefore, ignoring it would lead to upward biased estimation of covariance components.
Conversely, Schutz et al.11 estimated direct maternal genetic covariance for 305-day MY being lower (138.660 kg) than those obtained in the current study in first, second and third lactations. Negative direct-maternal genetic covariance suggests that selection for short days open will be associated by correlated increase in milk yield. Regarding fat yield, direct maternal genetic covariances were lower than (-14 kg) those obtained in the present study.
Diop et al.26 applied multiple-trait animal model by REML using three models to estimate direct, maternal, grand maternal genetic variances for growth traits in Gobra beef cattle and they concluded that an efficient model should include direct genetic, maternal genetic and direct-maternal genetic covariances that maternal genetic effects are essential specially at 18 months of age and they argued that grand maternal effect could be excluded from the model.
Conflicting results were recorded by Schutz et al.11 who reported that maternal genetic effects are not important in most of yield traits in dairy cattle but only they can improve the accuracy of animal model. Furthermore, they stated that direct additive genetic and maternal genetic effects did not affect phenotypic variance of milk yield, fat yield and fat percentage for Holsteins in New York.
El-Awady et al.27 estimated variance, covariance, genetic parameters and breeding values using multi-trait animal model (MTDFREML) and concluded that presence of maternal effect associated by presence of genetic variation which enables mass selection for growth traits in Rahmani lambs in Egypt. They added maternal effect should be considered in breeding value estimation that excluding maternal effect during breeding value estimation may lead to inefficient breeding value. Different from present findings, Hassen et al.28 pointed out that direct additive genetic variances and direct heritability estimates were higher than maternal genetic variances and maternal heritability estimates for early growth traits in lambs.
Referring to heritability, Konig et al.24 calculated direct heritability using VCE-REML for 305-day MY (0.35±0.028) and DO (0.03±0.004) to be smaller than those present in the current study except models 1 and 3 in second lactation and model 3 in third lactation for milk yield and model 1 in second lactation for days open. Increasing heritability in model 2 means that when the maternal effects are present but not considered the estimate of direct heritability would decrease than if maternal genetic effects are considered. Moreover, Mohammadi et al.29 argued that low direct genetic association between all traits indicated low genetic response would be expect through mass selection and indirect selection should be applied such as reproductive traits for improvement of growth traits in shal sheep.
Concerning breeding value, Sahin et al.30 observed that breeding values for 305-day MY were positive in years and negative in others, which may be due to unknown reasons or using sires with unknown estimated breeding values. They concluded that breeding value has a great role in genetic improvement of production traits. However, Tilki et al.31 mentioned that there is no positive or negative breeding values observed during different years.
Standard deviations of the present study showed high estimates in different models in all lactations. High standard deviations for these traits indicate the presence of high genetic variations among animals and eventually increase the possibility of selection of sire for these traits, which reduce bias from selection and achieve the best accuracy of predictions. The previous findings are in consonance with the findings of Kadarmideen et al.18 and Shalaby et al.20.
Regarding the spearman correlation coefficients of the breeding values, Shokrollahi and Baneh32 stated that if there was a positive genetic correlation between two traits this indicates that the same genes or genes with pleiotropic functions influence these traits and selection policy for these traits should depend on selection for these traits at each stage of life such as weight gain in later ages leads to improvement of body weight at older age in Arabi sheep.
CONCLUSION
The present study contributes to the comparison of three models to estimate the variance components, genetic parameters, breeding values and spearman genetic correlation coefficients of six productive traits affecting milk yield. The present results indicate the importance of presence of maternal genetic effect during planning selecting strategies especially if the variation between animals due to maternal effect is high.
REFERENCES
- Berry, D.P., P. Lonergan, S.T. Butler, A.R. Cromie, T. Fair, F. Mossa and A.C.O. Evans, 2008. Negative influence of high maternal milk production before and after conception on offspring survival and milk production in dairy cattle. J. Dairy Sci., 91: 329-337.
CrossRefDirect Link - Gudex, B.W., D.L. Johnson and K. Singh, 2012. Prenatal maternal effects on daughter milk production in New Zealand Dairy cattle. Proc. N. Z. Soc. Anim. Prod., 72: 19-22.
Direct Link - Legates, J.E., 1972. The role of maternal effects in animal breeding: IV. Maternal effects in laboratory species. J. Anim. Sci., 35: 1294-1302.
PubMedDirect Link - Willham, R.L., 1963. The covariance between relatives for characters composed of components contributed by related individuals. Biometrics, 19: 18-27.
CrossRefDirect Link - Pantelic, V., L. Sretenovic, D. Ostojic-Andric, S. Trivunovic, M.M. Petrovic, S. Aleksic and D. Ruzic-Muslic, 2011. Heritability and genetic correlation of production and reproduction traits of Simmental cows. Afr. J. Biotechnol., 10: 7117-7121.
Direct Link - Robison, O.W., 1981. The influence of maternal effects on the efficiency of selection: A review. Livest. Prod. Sci., 8: 121-137.
CrossRefDirect Link - Jurado, J.J., A. Alonso and R. Alenda, 1994. Selection response for growth in a Spanish Merino flock. J. Anim. Sci., 72: 1433-1440.
PubMedDirect Link - Seykora, A.J. and B.T. McDaniel, 1983. Heritabilities and correlations of lactation yields and fertility for holsteins. J. Dairy Sci., 66: 1486-1493.
CrossRefPubMedDirect Link - Visscher, P.M. and R. Thompson, 1992. Comparisons between genetic variances estimated from different types of relatives in dairy cattle. Anim. Prod., 55: 315-320.
CrossRefDirect Link - Schutz, M.M., A.E. Freeman, D.C. Beitz and J.E. Mayfield, 1992. The importance of maternal lineage on milk yield traits of dairy cattle. J. Dairy Sci., 75: 1331-1341.
CrossRefDirect Link - Huizinga, H.A., S. Korver, B.T. McDaniel, and R.D Politiek 1986. Maternal effects due to cytoplasmic inheritance in dairy cattle. Influence on milk production and reproduction traits. J. Livest. Prod. Sci 15 : 11-26.
CrossRefDirect Link - El Fadili, M., C. Michaux, J. Detilleux and P.L. Leroy, 2000. Genetic parameters for growth traits of the Moroccan Timahdit breed of sheep. Small Rumin. Res., 37: 203-208.
CrossRefDirect Link - Robertson, A., 1959. The sampling variance of the genetic correlation coefficient. Biometrics, 15: 469-485.
CrossRefDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Kadarmideen, H.N., R. Thompson, M.P. Coffey and M.A. Kossaibati, 2003. Genetic parameters and evaluations from single- and multiple-trait analysis of dairy cow fertility and milk production. Livest. Prod. Sci., 81: 183-195.
CrossRefDirect Link - Shalaby, N.A., M.A. Mostafa, M.F. Abdel-Glil and M.A.Z. Aleman, 2009. Genetic evaluation for milk production, reproduction and somatic cell count traits in early lactation for Friesian cattle in Egypt. Mansoura Univ. J. Agric. Sci., 34: 9463-9475.
Direct Link - Hussain, A., J. Iqbal, M. Shafiq, P. Akhtar, A. Shakoor, U. Waheed and S. Campus-Jhang, 2014. Genetic and phenotypic trends for milk yield per lactation in sahiwal cattle under arid and semi arid conditions of Pakistan. Pak. J. Life Soc. Sci., 12: 170-173.
Direct Link - Kadarmideen, H.N., R. Thompson and G. Simm, 2000. Linear and threshold model genetic parameters for disease, fertility and milk production in dairy cattle. Anim. Sci., 71: 411-419.
Direct Link - Konig, S., N. Chongkasikit and H.J. Langholz, 2005. Estimation of variance components for production and fertility traits in Northern Thai dairy cattle to define optimal breeding strategies. Archiv Tierzucht, 48: 233-246.
Direct Link - Meyer, K., 1992. Variance components due to direct and maternal effects for growth traits of Australian beef cattle. Livest. Prod. Sci., 31: 179-204.
Direct Link - Diop, M., J. Dodenhoff and L.D. Van Vleck, 1999. Estimates of direct, maternal and grandmaternal genetic effects for growth traits in Gobra cattle. Genet. Mol. Biol., 22: 363-367.
Direct Link - El-Awady, H.G., E.Z.M. Oudah, N.A. Shalaby, M.N. El-Arian and H.R. Metawi, 2011. Genetic improvement study on pre-weaning body weight of Egyptian Rahmani lambs under a pure breeding production system. Options Mediterraneennes, 100: 311-316.
Direct Link - Hassen, Y., B. Fuerst-Waltl and J. Solkner, 2003. Genetic parameter estimates for birth weight, weaning weight and average daily gain in pure and crossbred sheep in Ethiopia. J. Anim. Breed. Genet., 120: 29-38.
CrossRefDirect Link - Mohammadi, H., M.M. Shahrebabak, H.M. Shahrebabak, A. Bahrami and M. Dorostkar, 2013. Model comparisons and genetic parameter estimates of growth and the Kleiber ratio in Shal sheep. Archiv Tierzucht, 56: 264-275.
Direct Link - Sahin, A., Z. Ulutas, A.Y. Adkinson and R.W. Adkinson, 2014. Genetic parameters of first lactation milk yield and fertility traits in Brown Swiss cattle. Ann. Anim. Sci., 14: 545-557.
CrossRefDirect Link - Tilki, M., M. Colak and M. Sari, 2009. Genetic parameters of 305-day milk yield for brown Swiss reared in the bahri dagdas international agricultural research institute in Turkey. Kafkas Univ. Vet. Fak Derg., 15: 397-400.
CrossRefDirect Link - Shokrollahi, B. and H. Baneh, 2011. (Co)variance components and genetic parameters for growth traits in Arabi sheep using different animal models. Genet. Mol. Res., 11: 305-314.
CrossRefDirect Link