
Sermin Tagil
Department of Geography, Faculty of Arts and Sciences, Balikesir University, 10100, Balikesir, Turkey
Journal of Applied Sciences
Year: 2015 | Volume: 15 | Issue: 6 | Page No.: 850-861







ABSTRACT
The analysis of habitat factors for the distribution of landuse-landcover (LULC) which is based on the analysis of Topographic Habitat Characteristics (THC), is important to understand the process of island ecology. The relationships between LULC, especially relative abundances of dominant tree covers and THC were examined in the Kapidag Peninsula in order to understand the spatial heterogeneity of LULC. Dominant tree covers and LULC patch mosaics were obtained from Turkish General Directory of Forestry (GDF) by using categorical dataSeveral THC, indicating temperature and moisture distribution, exposure to wind etc., were computed by a 15 m resolution Digital Elevation Model (DEM). Simple chi-square test and contingency tables were used to explore the connections between LULC types and THC. Landform description has long been recognized essential for understanding vegetation distribution in the peninsula. In this study, the lack of information on the human dimension is the reason of some uncertainties in interpreting the spatial patterns of vegetation, in the peninsula.
PDF Abstract XML References Citation
Received: January 22, 2015;
Accepted: April 03, 2015;
Published: May 23, 2015
How to cite this article
Sermin Tagil, 2015. Effect of Topographic Habitat Characteristics on the Spatial Distribution of Landuse-Landcover in the Kapidag Peninsula, Turkey. Journal of Applied Sciences, 15: 850-861.
DOI: 10.3923/jas.2015.850.861
URL: https://scialert.net/abstract/?doi=jas.2015.850.861
DOI: 10.3923/jas.2015.850.861
URL: https://scialert.net/abstract/?doi=jas.2015.850.861
INTRODUCTION
Information related to the characteristics and spatial distribution of vegetated and unvegetated land cover is critical for monitoring and managing the environment. Much of the environmental variability attributed to topographical factors, controls the spatial patterns of landcover (Gemici and Secmen, 1990; Brown, 1994; Del Barrio et al., 1997). The relationships between ecosystem composition and topographic features based on precipitation and solar radiation in different ecosystems have been widely studied (Vankat, 1982; Gemici and Secmen, 1990; Moore et al., 1993b; Pinder et al., 1997; Ozel, 1999; Oner, 2010). Most of these studies have focused on spatial patterns of the vegetation.
Although landforms are relatively stable landscape features, they are important input parameters for spatial modeling of vegetation distribution (Hoersch et al., 2002). Thus, it can be said that topography is the reason of patchwork pattern of small scale habitats (Hoersch et al., 2002), because species-environment relationships are important determinants of the abundance and spatial distribution of vegetation. The first quantification of landform characteristics to predict vegetation distribution was done by Parker who developed a topographic relative moisture index (Parker, 1982, 1989). Most of the studies were done about the effect of topography on vegetation composition (Gemici and Secmen, 1990; Bolstad et al., 1998; Franklin, 1995; Deng et al., 2007a; Ozel, 1999; Oner, 2010) and species abundance (Gemici and Secmen, 1990; Meentemeyer et al., 2001). According to Horsch (2003), the distribution of actual vegetation is not only a result of the complex interaction of historical and recent environmental factors but also of humane and disturbance factors.
The rich array of topographic data and digital terrain analysis techniques have advantage to show the effect of topographic habitat characteristics on spatial distribution of land-cover (Deng et al., 2007b; Wilson and Gallant, 2000). Because Digital Elevation Model (DEM) highly correlates with moisture, temperature, geomorphologic processes it is a potential substitute for use in vegetation analysis (Franklin et al., 2000; Moore et al., 1991; Hoersch et al., 2002). Digital vegetation information as well as landform and topography-dependent climatic conditions are commonly assessed using Geographic Information Systems (GIS).
Topographical characteristics have importance on water availability. The effects of microtopoclimatic contrasts appear more important in regions where general aridity promotes critical variation in moisture content (Churchill, 1981; Campbell, 1989). Because of this, topographical characteristics are important factors in shaping Mediterranean vegetation patterns (Armesto and Martinez, 1978; Ozel, 1999; Oner, 2010). The rates of the changes in water-limited Mediterranean vegetation would be influenced by topographic conditions.
In this study, the influence of certain THC on the spatial patterns, not only of vegetated patches but also of un-vegetated patches, was studied in the Kapidag Peninsula located in the Mediterranean biological climate. This study aims to describe LULC types and their spatial distribution patterns in the peninsula, to check whether the spatial distribution of LULC types is controlled by topographic attributes and to examine how specific THC influence spatial heterogeneity in this landscape.
When the spatial distribution of the main land-cover types and their relations with topographical factors were statistically analyzed, GIS-based model was applied. In this study, we do not have information on the human dimension. Also, biotic processes driving landscape patchiness are outside the scope of this study.
MATERIALS AND METHODS
Study site (area of research and characteristics for vegetation distribution: The present Kapidag Peninsula (Peninsula of Cyzicus) was formerly an island and later became attached to land as a tombolo (Erol, 1991). The Kapidag peninsula is nearly 300 km2 and is in the north of Balikesir province, it lies between 27°30 W-28°15 E longitude and 41°15 N-41°43 S latitude (Fig. 1). Elevations in the study area range from 0 m (0 ft) to 805 m (2641 ft). Kapidag Peninsula (Arktonnesos) is linked to Turkey mainland by the Belkis tombolo (Isthmus). It is a large (1500 m in length along its central axis and 1700 m wide) sand tombolo marked by alluvial flat land (Ardel and Inandik, 1957). Kapidag peninsula, bends the incoming waves around it so that their energy sweeps sand onto the Belkis tombolo from both sides. This area is different from the surrounding mountainous areas in terms of vegetation cover.
Cayagzi, Erdek, Ilhankoy, Karsiyaka, Ocaklar, Ormanli, Sahinburgaz, Tatlisu and Turankoy are important settlements in the peninsula. Most people in the area are engaged in agricultural activities dealing with vineyards, fruit orchards and olive plantations.
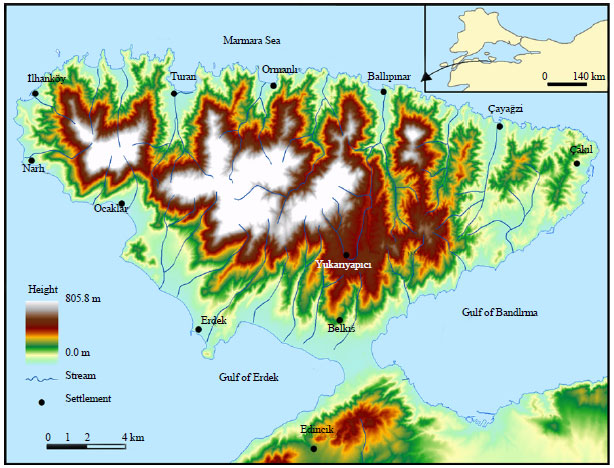 | |
| Fig. 1: | Area of research |
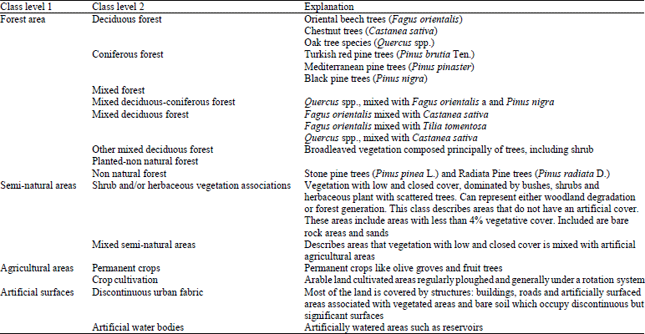
| Table 1: | Hierarchical classification scheme |
 | |
In the study area, the annual precipitation is 695 mm and the annual average temperature is 12.4°C (Erdek meteorology stations). According to Thornthwaite (1948), the study area was classified as C2b2 s2b3 which is semi humid, second-degree mesothermal, under a sea climate effect and has a strong water deficit during summertime. Generally, climate is a semi-humid mild type (Sonmez, 2001).
Because Kapidag peninsula has a more humid climate than its surrounding lands, it has wealthier flora. Three main vegetation types are found in the study area: Humid forest, dry forest and macchie. As Sonmez (2001), the distribution rates of the taxa into phytogeographical regions are as follows: Euro-Siberian (57%), Mediterranean (26%) and Irano-Tuanian (17%). Most common forest species (Tasturen, 1994; Sonmez, 2001) are oriental beech trees (Fagusorientalis lipsky), chestnut trees (Castanea sativa Mill.), lime trees (Tiliaplaty phyllos Scop.), oak tree species (Quercus spp.), Turkish red pine trees (Pinus brutia Ten.), Mediterranean pine trees (Pinus pinaster Ait) and black pine trees (Pinus nigra Var.). Dominated oak species are Quercus cerris L., Q. coccifera L., Q. frainetto Ten., Q. infectoria Olivier, Q. petrea (Mattuschka) Liebl., Q. robur L.
Spatial data:
Dependent variables (ground cover): We created dominant tree covers and LULC patch mosaics using categorical data obtained from Turkish General Directory of Forestry (GDF). LULC patch mosaics are based on the dominant human landuse and the dominant vegetation forms. Dominant tree species are based on stand types distinguished on the basis of crown closure, stand development stage and tree species. From 295 stand types, 4 main land cover classes and 18 sub-classes were done (Table 1). While sub-classifying, dominant tree species were taken into consideration. In both cases, the original data in vector format was converted to raster data sets using a 15 m size cell. Classification was used to group stands with similar species composition in order to simplify the analysis process of landscape-level vegetation.
Predictor variables (dem and topographic habitat characteristics): Habitats can be dramatically different depending on Topographic Habitat Characteristics (THC). DEMs offer many more potential habitat descriptors than simply a set of elevation values (Tagil and Jenness, 2008). A 20 m resolution DEM was computed from 10 m contours of a 1:25000 topographic map. Different topographic-geomorphometric parameters can be derived from the DEM by using GIS approaches.
A ten terrain-based topographic habitat models derived from the DEM to determinate the distribution of plants and ecological communities was created. THC derived and their indication potential are given in Table 2. Firstly, 8 landform variables were created besides primary parameters like elevation; secondary parameters (slope position, landforms, etc.) were also created by combining primary ones. These models were used in a wide variety of ecological applications (Newell and Peet, 1998; Anderson et al., 1998; Franklin et al., 2000). The THC used in the study represent light related variables, water availability, temperature, micro-climate and landscape unit, because there was not any spatial data about temperature, moisture, wind or snow in the Kapidag Peninsula. However, we know that landform derivatives are to be used as substitutes (Moore et al., 1991; Hoersch et al., 2002).
| Table 2: | Terrain-based models created from DEM at 15 m resolution |
 | |
Sampling design and data analysis: We selected 1046 sample locations at random all over the study area maintaining a minimum distance of 500 m between sample points. Random sampling technique is often used to eliminate spatial dependency (Wagner and Fortin, 2005). Initial analysis was made on correlations between LULC types and individual topographic habitat variables.
Analysis of correlation between vegetation and landform variables: THC and LULC variables were uniformly scaled to calculate the statistical correlations between the response and predictor. The only appropriate method to analyze correlations is the contingency table for non-metric factor data, because the LULC data is on the nominal scale. Due to this, the topographic habitat characteristics were transferred into nominal data. Therefore, classes were used instead of raw values (Table 2). Presence/absence of all LULC types was considered for each sample points.
Because of nonparametric tests’ distribution, normal distribution of the population was not tested from which the data are drawn. After variables were classified, the statistical significance of the relationships between each THC and the abundance of LULC types was determined using χ2 (Chi square) contingency tables (Steel and Torrie, 1980). For analysis, χ2 contingency tables were built up for every one of the LULC classes as shown in Table 1 and every one of the THC, as shown in Table 2 to test the null hypothesis of no connection between LULC types and terrain attribute.
Analyses for all topographic habitat characteristics with all LULC classes: Firstly the relations between single topographic habitat characteristics and LULC level 1 classes were investigated, secondly more-dimensional relationships were focused on by using LULC level 2 data (Table 1). The analysis was performed using GIS summarizing and filtering tools. All statistical analyses were conducted using SPSS software.
RESULTS
Climatic factors such as temperature and rainfall are the direct factors of a physiological influence on species but they are not topographic variables (Austin and Smith, 1990). Although the use of direct factors is preferable for predicting biodiversity (Coblentz and Riitters, 2004), this kind of data may not be available, particularly for large regions. Because of this, in this study, the spatial differences of LULC based on topographic parameters which were readily measured were explored . DEM derived from measures of slope, aspect, TWI, solar insolation, rugosity, slope position and landforms were used to represent the climatic elements which are potentially important for LULC and also vegetation distribution. The distributions of topographic characteristics are presented in Fig. 2, respectively.
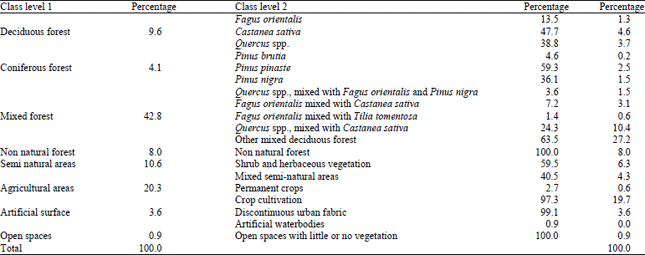
Forest lands are the major land cover type, covering 56.6% of the study area; this was followed by crop cultivation areas (20.3%) and semi natural areas (10.6%; Table 1). Most (42.8%) of the total forest land are covered by mixed forest land (Table 3).
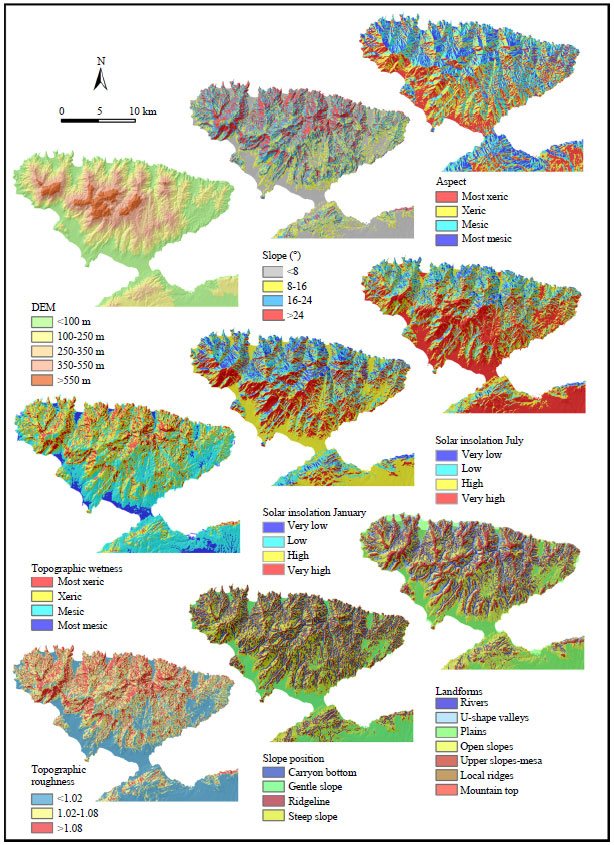 | |
| Fig. 2: | Distributions of topographic habitat characteristics in the Kapidag Peninsula |
Oriental beech (Fagus sylvatica) and Oak (Quercus sp.) forests are the most common deciduous forest lands. Stone pine (Pinuspinea) are the most abundant coniferous forest in the study area, because it has been so widely planted throughout the Mediterranean region. Artificial surfaces and open spaces have little coverage.
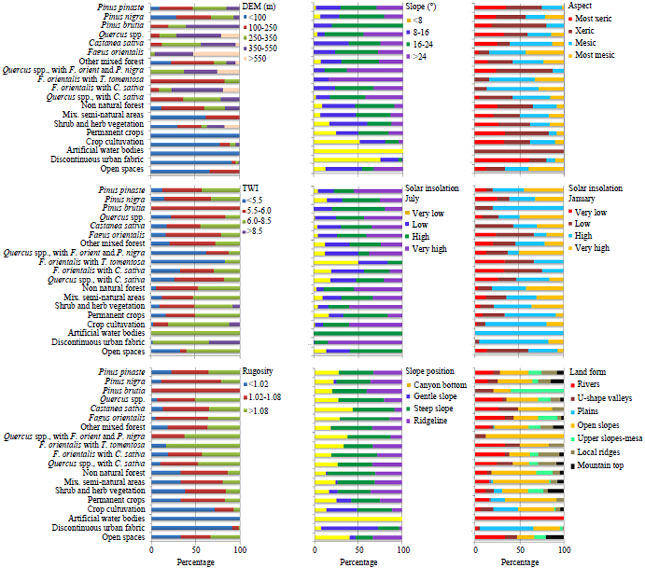 | |
| Fig. 3: | Associations between types of LULC and topographic characteristics |
Chi-square test has been performed to test the homogeneity of intraclass correlations of landcover classes and significant difference was found between LULC in both classification levels (p<0.0001).
To demonstrate whether the spatial distribution of LULC types is associated with topographic characteristics, a contingency table was built up and χ2 test was performed. Chi-square was used to test the null hypothesis that the disturbance of LULC in both classification levels on landscape differ significantly from one another with respect to how well they are linked We reject the null hypothesis of no association and conclude that there is a relationship between LULC in level 1 and THC (p<0.001). Again the relation between THC and each LULC at the level 2 was assessed using cross classification technique and Pearson’s Chi-squared test. Figure 3 shows the deviations in frequencies observed with regard to those estimated during the χ2 test and makes it possible to reveal specific associations between types of LULC and THC.
First of all elevation and LULC-level 2 types will be discussed, because the elevation determines the distribution of vegetation and is a typical characteristic of mountain regions. The natural distribution of forests is sensitive to elevation because of the physiological requirement of plant species. In the peninsula, elevation ranged from 0-805.8 m. The altitudinal ranges were categorized into five classes. The elevation category of <100 m is the most dominant (36.3%), followed by the altitudinal ranges of 100-250 m (30%). Only about 13.8 and 14.0% of the areas are at the altitudinal range of 250-350 and 350-550 m, respectively. The altitude higher from 550 m has the lowest cover (5.9%). The overall Pearson Chi-square revealed a significant association between LULC and elevation, p<0.001. Therefore, it can be deduced that the distribution of LULC is strongly correlated with elevation. However, in detailed thematic scale (LULC level 2), relationship is slightly more complex (Table 4). We can say that the distribution of LULC types is related to elevation and most of these relationships are statistically significant (Table 4).
| Table 3: | Frequency distribution of a LULC variables |
 | |
| Table 4: | Summary of the Pearson’s Chi-square test for categorical variables |
 | |
| *Significant level of 0.05, **Significant level of 0.01 | |
This shows that elevation is controlled the presence/absence of almost all LULC classes at level 2. However, predictor variable "elevation" was not significantly associated with coniferous forest Pinus brutia and Pinus nigra and artificial water bodies at a significant level of 0.05. The upper altitudes of the peninsula (>550 m) supports the optimum distribution of Fagus orientalis, because this altitudinal range is characterized by northerly winds all year round and byan increase in cloudiness with altitude and annual rainfall maximum. For example, as seen on Fig. 3, 52.4% of Fagus orientalis prefer altitude higher than 550 m, compared to 20% of Quercus spp. And 4.3 % of Castanea sativa prefer this altitude. Fagus orientalis were almost twice as likely to prefer the highest altitude in the peninsula. A strong positive change in proportional abundance of Fagus orientalis was found with increasing elevation (Fig. 3). For the rest of the LULC types, the proportional area they occupy decreases with higher elevations. The proportion of anthropogenic land covers becomes higher with declining elevations (Fig. 3).
Slope is another important predictor variable. In the area, it ranges from 0- 45°. The slope category of range of 16-24° is the most dominant (31.3%), followed by the slope lower than 8° (25.1%). About 24.5 and 19.2% of the areas are at the slope range of 8-16 and >24°, respectively (Fig. 2). All LULC categories exhibited more or less ball-shaped distributions along the slope categories (Fig. 3). Slope was significantly associated with deciduous forest Castanea sativa and Quercus spp., coniferous forest Pinus pinaster, all type of mixed forests, non natural forest and shrublands at a significant level of 0.01 and with crop cultivation and urban fabrics at a significant level of 0.05 (Table 4). A decrease in the relative abundance of coniferous forest, shruplands and anthropogenic land covers (Urban fabric, water bodies, crop cultivation) can be observed with increased slope angle. Moreover, an increase in the relative abundance of mixed forests can be observed with increased slope angle, except for oriental beech trees mixed with chestnut trees (Fig. 3). Steeper topographic features are not suitable for crop production due to inaccessibility and fragility of the land. Because of this, the high proportion of crop cultivation and the low forest cover on gentle slopes are apparently the results of anthropogenic activities. Two groups of land cover classes, namely agriculture and discontinuous urban fabric, were mainly found at gentle slopes (<8°) while shrubland and forest lands had their dominant existence at steeper slopes. But in the peninsula, as it is also revealed by the results shown in Fig. 3, there are steep slopes which are used for permanent crop due to scarcity of land. Shortly, steeper slopes favor the survival of forests.
The topographic variable aspect influences parameters such as exposure to sunlight, drying winds and evapo-transpiration. Therefore, aspect has implications to physiological and ecological requirements of the species. Several studies have been done to show the effect of aspect on separation of plant communities (McNab et al., 1999; Ozel, 1999; Zhang et al., 2006; Oner, 2010). In the study, an aspect map of the peninsula was produced showing four thematic categories (Table 2). Mesic aspect is the most dominant (31.12%), followed by the most xeric (25.13%) and xeric (24.54%) aspects. Most mesic aspects has the lowest cover (19,21%). There is significant difference between LULC level 1 in terms of which type of aspect they are on (p<0.001). In detailed thematic scale (level 2), an association between aspect and Castanea sativa, Fagus orientalis mixed with Castanea sativa and urban fabric are found, p<0.001 and also an association between aspect and Pinus pinaster, Quercus species mixed with Castanea sativa, non natural forest and crop cultivation are found, p<0.05 (Table 4). It can be hypothesized that presence/absence of the Castanea sativa and Fagus orientalis forests are controlled by aspect, more than another type of LULC. 42% of Fagus orientalis prefers most mesic aspects and 48% of Castanea sativa and 42% of Quercus spp. species mixed with Castanea sativa prefer mesic aspects. The 40% of Pinus pinaster prefers xeric aspects. Most of the deciduous forest follows the normal pattern in the region, preferring mesic aspects. The agricultural land cover class peaks in the most xeric aspects while the highest occurrence of permanent crop is recorded in the xeric aspects. The differences between mesic and xeric aspects could be because of water availability which cannot be explained by differential insolation. Mesic slopes have little sunlight and tend to be cooler than the south, where sun is predominantly shining. Species on the southern slopes are relatively more adapted to evaporation, hot and dry conditions while species on the north and northwest facing slopes need more water and cooler conditions. This knowledge could be instrumental in future efforts of species selection for plantation development in the study area.
The TWI indicates locations where water flows gravitationally down, ensuring higher surface wetness. The TWI has been used to characterize biological processes such as forest site quality, annual production and vegetation patterns (Moore et al., 1993b; White and Running, 1994). TWI has been useful for predicting the spatial distribution of species richness (Zinko et al., 2005; Sorensen et al., 2006). In the study, a TWI map of the peninsula was produced showing four thematic categories (Table 2). Mesic aspect is the most dominant (46.3%), followed by the xeric (34.6%) and most xeric (13.4%). Most mesic surface has little coverage with 5,7% (Fig. 3). TWI control presence/absence of some LULC classes but not all of them (Table 4). An association between TWI and Castanea sativa, Quercus spp., Quercus spp. mixed with Fagus orientalis and Pinus nigra, Fagus orientalis mixed with Tiliato mentosa, Quercus spp. mixed with Castanea sativa, other mixed forest, non natural forest, crop cultivation was found at a significant level of 0.01; with Pinus brutia and Fagus orientalis mixed with Castanea sativa at a significant level of 0.05. Presence/absence of the most of the deciduous forests was controlled by TWI.
Topography creates spatial variability of insolation which in turn generates variation in microclimate conditions such as temperature, humidity, snow cover and soil moisture. In mountainous regions; altitudinal gradients in temperature are also significant controls on LULC patterns (Hoersch et al., 2002; Deng et al., 2007a). We calculated the solar insolation for summer and winter by using DEM. Summer insolation is controlled by the presence/absence of the LULC classes more than winter insolation (Table 3). An association between Castanea sativa, Fagus orientalis and Quercus spp. mixed with Castanea sativa and Fagus orientalis mixed with Castanea sativa and July insolation is statistically significant at the level of 0.01. Conifer forest Pinus pinaster and Pinus nigra and also shruplands area associated with July insolation at the level of 0.05. More than these LULC types, July insolation controlled the presence/absence of the Urbanfabric. When winter insolation was examined, only Fagus orientalis mixed with Castanea sativa, Quercus spp. mixed with Castanea sativa, non natural forest, shrupland, crop cultivation and urban fabric had association. About 84% of the discontinuous urban fabric prefers very high solar insolation during the summer (Fig. 3) and 79% of it prefers high solar insolation in winter times. This means that sunny aspects are chosen for urbanization. It can be said that correlation between presence or absence of most of the LULC and the winter insolation is not perfect. Shortly, un-vegetated surfaces increase and vegetated surfaces decrease with potential daily short wave radiation (Fig. 3). It can be said that aspect is a poor indicator of received radiation because of the complex topography in the peninsula.
Rugosity, one of the topographic parameters of the landscape, can be readily measured from digital terrain data and in many settings they influence the distribution of landcover and landcover diversity (Coblentz and Riitters, 2004; Coblentz and Keating, 2008; Istanbulluoglu et al., 2008). In the study, landscape rugosity refers to the unevenness of the earth’s surface due to topography. Rugosity may be defined as the roughness of the physical structure of the peninsula surface. According to Chrysoulakis et al. (2004), rugosity affects transport of hydrometeorological fluxes between land surface and the atmosphere, as well as below the surface, i.e. infiltration and water movement. Because of this, rugosity controls presence/absence of the non natural areas like urban fabric, crop cultivation and non natural forest areas (Table 4). Also, rugosity is associated with Quercus spp. mixed with Castanea sativa, Pinus nigra, Quercus spp. and Castanea sativa at of the level of 0.01 but with Pinus brutia and Quercus spp. mixed with Fagus orientalis and Pinus nigra at of the level of 0.05. It demonstrates that there is a strong spatial correlation between areas of high topographic roughness and Fagus orientalis mixed with Tiliato mentosa (83%), Quercus spp. mixed with Castanea sativa and Pinus nigra (63%), Quercus spp. (51%), Quercus spp. mixed with Castanea sativa (47%) and Fagus orientalis mixed with Castanea sativa (42%). The 68% of Pinus nigra and 100% of the Pinus brutia prefer middle rugosity (Fig. 3). In the absence of data on habitat types, rugosity could be a useful surrogate for this habitats. Lower rugosity supports mainly crop cultivation and discontinuous urban fabric. About 91% of the discontinuous urban fabric and 71% of crop cultivation have been shown to be related to low rugosity. We find that there is a noticeable relationship between organization of some LULC types in the peninsula and the topographic roughness.
Slope position was classified as canyon bottom, gentle slope, steep slope and ridgeline. Steep slopes are the most dominant (41.4%), followed by the ridgeline (21.9%), canyon bottom (19.6%) and gentle slopes with 17.70% (Fig. 2). Castanea sativa, Fagus orientalis and crop cultivation are highly associated with slope position (p<0.001; Table 4). The 47 and 43% of Castanea sativa prefers respectively steep slopes and canyon bottoms. The 57% of Fagus orientalis have been shown on steep slopes and 47 and 43% of crop cultivation have been shown on the steep slopes and gentle slope (Fig. 3). Relationship with Quercus spp., Pinus pinaster, Quercus spp. mixed with Castanea sativa, shrub and her. veg. and artificial water bodies was found at a significant level of 0.01.
Terrain attributes were yielded in eight landforms. Figure 2 shows the spatial distribution of landforms. A Chi-square test result indicates the association of spatial distribution of LULC types and landforms (p<0,001). Landforms are controlled by the presence/absence of almost all of the LULC classes (Fig. 3). Predictor variable "landform" was highly associated with Castanea sativa, Quercus spp. mixed with Castanea sativa, other mixed forests, non natural forests, all semi natural areas, crop cultivation, urban fabrics (p<0.001, Table 4). Also, an association between landforms and all type of deciduous forests, all type of semi natural areas, crop cultivation, Pinus brutia, Fagus orientalis mixed with Castanea sativa, Quercus spp. mixed with Castanea sativa, other mixed forests, urban fabric are found, p<0.05. For example, 60% of the urban fabrics are on the plains; 68% of the crop cultivation are on the plains and open slopes. The 65% of the Castanea sativa areas are on the rivers and open slopes (Fig. 3). These trees grow where the soil moisture is very high (Atalay, 1992).
DISCUSSION
This study highlights the relationship between the topographic parameters and the spatial distribution of different LULC types in the Kapidag Peninsula. Elevation, slope, aspect, TWI, solar insolation, rugosity, slope position and landform index, that quantify topographic shape and protection, were used to represent the climatic elements and to distinguish landforms and predict ecosystem distribution on the Kapidag Peninsula. In this study, the hypothesis that topographic parameters exert an important control on the spatial LULC distribution was tested.
The spatial analyses of land cover in relation to environmental variables indicated the overall situation of the peninsula. A Chi-square test result demonstrates the association of spatial distribution of LULC types and the landform parameters (elevation, slope, aspect, radiation, etc.). In a general perspective, the spatial distribution of LULC of Kapidag Peninsula is affected by the THC. Different terrain characteristics have different roles on LULC distribution. For example, different landover classes concentrate on different aspects.
When it is examined in detail, tree species’ abundance and distribution patterns in the peninsula are correlated with terrain characteristics and indirectly, climate and also potential soil moisture gradients. All of the THC significantly predict species abundance or composition. It depends on the ecophysiological tolerance of tree species to drought (Linton et al., 1998). Species (Pinus brutia, Pinus pinaster, etc.) tolerant of drought dominate lower elevations while more mesophytic species (Fagus orientalis, Castanea sativa, etc.) dominate moist, high elevation sites. Studies on tree species water relations in the peninsula could provide a mechanistic explanation for the observed abundance and distribution patterns of vegetation in peninsula. This study also indicated that forest ecosystem is found in the high altitude and steep areas of the peninsula. Generally, agriculture is scarce on gentle terrain while forest lands develop preferentially on high and steep terrain. However, other influence factors have to be kept in mind, especially the human factor. This factor cause diverse spatial patterns, being somehow different from a natural pattern. Although, there is no spatially explicit information on the human dimension, THC derived from a DEM are capable of explaining a great amount of the spatial variability of LULC alliances within the study area, because relationships between almost all LULC classes and THC are found statistically significant at the level of 0.01. Major anthropogenic activities (crop cultivation and urbanization) are dominantly undertaken on the gentle slopes, low altitudes, southern slopes, plains. Major ecosystem on this area has changed by human activities and natural ecosystems are turning into crop cultivation and other human activities.
The study of LULC distribution cannot be easily explained by the results of this study, because of that there are many added elements of complexity, changes in landuse practices, site productivity and soil composition, etc. Prediction of distribution based on site variables works well for some LULC classes and is more difficult for others, in the area. Local scale interactions affected some LULC distributions more strongly than others. However, present results compared to those of other studies, ultimately influence which species are found at a specific site within the peninsula. Oner (2010) studied forest composition in the same area, comparing elevation, aspect, slope and landform with species composition. The results of this study also found elevation to be a significant predictor (Oner, 2010). Oner (2010) found that Fagus orientalis and Castanea sativa are most concentrated on northerly aspects. However, they state that Pinusbrutia Ten. and shruplands are most concentrated on southerly aspects. Also, Sonmez (2001) found similar results. Our study found that basal area and density of Fagus orientalis was most concentrated on most mesic, it means northeast aspects, Pinus brutia Ten. And shruplands basal area and density was most concentrated on Xeric-southwest aspects. Also, Oner (2010) gives some information about species’geomorphological choice. For example Oner (2010) found that Fagus orientalis distribution was most concentrated in steep cliff areas, similar to our findings. Like our study, Oner (2010) and Sonmez (2001) also note that human impacts, such as fire suppression and population growth, are affecting forest composition and intense human pressure should be considered. Like our study, some species followed clear distribution patterns, such as Fagus orientalis and Castanea sativa, while other species, such as Pinus brutia Ten., appeared to be randomly distributed with preposition certain attributes. Similar with present study, Alemdag (1963) and Atalay (1992), also indicate that Fagus orientalis are mostly above 500 m where the fog and precipitation increase.
The results also showed that GIS is effective in monitoring the overall status and analyzing land cover patterns as a function of environmental parameters such as elevation, slope and aspect. On the other hand, different habitat association test methods may produce different results. In this study, results show that contingency tables are useful methods to quantify the potential of topographic characteristics for the indication of LULC distribution.
This study can be used as a basis of information for further identification of different wildlife habitats, ecological conditions and biodiversity conservation and management. The land cover information existing in the peninsula helps to develop the carrying capacity of the important species. Results also show that the impact of topography on the spatial LULC patterns must be analyzed, because of that topography has a main control in most landscapes to understand the heterogeneous spatial distribution of LULC. Detection of different environmental parameters might also help to know, plan and create a diversity of habitats available to plant species and contribute to overall increase of the species diversity. Therefore, conservation sites and management units in the Kapidag Peninsula can be identified and mapped with the help of GIS to set up development and management strategies in relation to environmental parameters preferred by the respective wildlife.
The nature of this study results in some limitations on the accuracy of our data. While values for DEM reflect how shady sites are due to topography, they do not reflect shade created by the forest canopy. In addition, incident radiation values derived from McCune and Keon (2002) equations reflect the amount of radiation at a site without a canopy cover.
REFERENCES
- Armesto, J.J. and J.A. Martinez, 1978. Relations between vegetation structure and slope aspect in the mediterranean region of Chile. J. Ecol., 66: 881-889.
Direct Link - Austin, M.P. and T.M. Smith, 1990. A new model for the continuum concept. Adv. Veg. Sci., 11: 35-47.
CrossRefDirect Link - Brown, D.G., 1994. Predicting vegetation types at treeline using topography and biophysical disturbance variables. J. Veg. Sci., 5: 641-656.
CrossRefDirect Link - Bolstad, P.V., W. Swank and J. Vose, 1998. Predicting Southern Appalachian overstory vegetation with digital terrain data. Landsc. Ecol., 13: 271-283.
CrossRefDirect Link - Chrysoulakis, N., I. Keramitsoglou and C. Cartalis, 2004. Hydrologic land cover classification mapping at the local level with the combined use of ASTER multispectral imagery and GPS measurements. Proceedings of the SPIE 5239, Remote Sensing for Environmental Monitoring, GIS Applications and Geology III, September 8, 2003, Barcelona, Spain, pp: 532-541.
CrossRefDirect Link - Churchill, R.R., 1981. Aspect-related differences in badlands slope morphology. Ann. Assoc. Am. Geogr., 71: 374-388.
Direct Link - Coblentz, D. and P.L. Keating, 2008. Topographic controls on the distribution of tree islands in the high Andes of South-Western Ecuador. J. Biogeogr., 35: 2026-2038.
CrossRefDirect Link - Coblentz, D. and K.H. Riitters, 2004. Topographic controls on the regional-scale biodiversity of the south-western USA. J. Biogeogr., 31: 1125-1138.
CrossRefDirect Link - Del Barrio, G., B. Alvera, J. Puigdefabregas and C. Diez, 1997. Response of high mountain landscape to topographic variables: Central pyrenees. Landsc. Ecol., 12: 95-115.
CrossRefDirect Link - Deng, Y., X. Chen, E. Chuvieco, T. Warner and J.P. Wilson, 2007. Multi-scale linkages between topographic attributes and vegetation indices in a mountainous landscape. Remote Sens. Environ., 111: 122-134.
CrossRef - Deng, Y.X., J.P. Wilson and B.O. Bauer, 2007. DEM resolution dependencies of terrain attributes across a landscape. Int. J. Geogr. Inform. Sci., 21: 187-213.
CrossRefDirect Link - Franklin, J., P. McCullough and C. Gray, 2000. Terrain Variables used for Predictive Mapping of Vegetation Communities in Southern California. In: Terrain Analysis: Principles and Applications, Wilson, J.P. and J.C. Gallant (Eds.). John Wiley and Sons, New York, USA., ISBN-13: 9780471321880, pp: 331-354.
- Franklin, J., 1995. Predictive vegetation mapping: Geographic modelling of biospatial patterns in relation to environmental gradients. Prog. Phys. Geogr., 19: 474-499.
Direct Link - Horsch, B., 2003. Modelling the spatial distribution of montane and subalpine forests in the central Alps using digital elevation models. Ecol. Mod., 168: 267-282.
CrossRefDirect Link - Hoersch, B., G. Braun and U. Schmidt, 2002. Relation between landform and vegetation in alpine regions of Wallis, Switzerland. A multiscale remote sensing and GIS approach. Comput. Environ. Urban Syst., 26: 113-139.
CrossRefDirect Link - Istanbulluoglu, E., O. Yetemen, E.R. Vivoni, H.A. Gutierrez-Jurado and R.L. Bras, 2008. Eco-geomorphic implications of hillslope aspect: Inferences from analysis of landscape morphology in central New Mexico. Geophys. Res. Lett., Vol. 35.
CrossRefDirect Link - Linton, M.J., J.S. Sperry and D.G. Williams, 1998. Limits to water transport in Juniperus osteosperma and Pinus edulis: Implications for drought tolerance and regulation of transpiration. Funct. Ecol., 12: 906-911.
CrossRefDirect Link - McCune, B. and D. Keon, 2002. Equations for potential annual direct incident radiation and heat load. J. Veg. Sci., 13: 603-606.
CrossRefDirect Link - McNab, W.H., S.A. Browning, S.A. Simon and P.E. Fouts, 1999. An unconventional approach to ecosystem unit classification in Western North Carolina, USA. For. Ecol. Manage., 114: 405-420.
CrossRef - Meentemeyer, R.K., A. Moody and J. Franklin, 2001. Landscape-scale patterns of shrub-species abundance in California chaparral: The role of topographically mediated resource gradients. Plant Ecol., 156: 19-41.
CrossRefDirect Link - Moore, I.B., T.W. Norton and J.E. Williams, 1993. Modelling environmental heterogeneity in forested landscapes. J. Hydrolol., 150: 717-747.
CrossRefDirect Link - Moore, D.M., B.G. Lees and S.M. Davey, 1991. A new method for predicting vegetation distributions using decision tree analysis in a geographic information system. Environ. Manage., 15: 59-71.
CrossRefDirect Link - Newell, C.L. and R.K. Peet, 1998. Vegetation of linville gorge wilderness, North Carolina. Castanea, 63: 275-322.
Direct Link - Parker, A.J., 1989. Forest/environment relationships in Yosemite National Park, California, USA. Vegetatio, 82: 41-54.
CrossRefDirect Link - Parker, A.J., 1982. The topographic relative moisture index: An approach to soil-moisture assessment in mountain terrain. Phys. Geogr., 3: 160-168.
Direct Link - Pinder, III J.E., G.C. Kroh, J.D. White and A.M.B. May, 1997. The relationships between vegetation type and topography in Lassen Volcanic National Park. Plant Ecol., 131: 17-29.
CrossRefDirect Link - Tagil, S. and J. Jenness, 2008. GIS-based automated landform classification and topographic, landcover and geologic attributes of landforms around the Yazoren Polje, Turkey. J. Applied Sci., 8: 910-921.
CrossRefDirect Link - Thornthwaite, C.W., 1948. An approach toward a rational classification of climate. Geographical Rev., 38: 55-94.
CrossRefDirect Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Wagner, H.H. and M.J. Fortin, 2005. Spatial analysis of landscapes: Concepts and statistics. Ecology, 86: 1975-1987.
CrossRefDirect Link - White, J.D. and S.W. Running, 1994. Testing scale dependent assumptions in regional ecosystem simulations. J. Veg. Sci., 5: 687-702.
CrossRefDirect Link - Zhang, X., M. Wang, B. She and Y. Xiao, 2006. Quantitative classification and ordination of forest communities in Pangquangou National Nature Reserve. Acta Ecologica Sinica, 26: 754-761.
CrossRef - Zinko, U., J. Seibert, M. Dynesius and C. Nilsson, 2005. Plant species numbers predicted by a topography-based groundwater flow index. Ecosystems, 8: 430-441.
CrossRefDirect Link