
Jawdat Alebraheem
School of Mathematical Sciences, Universiti Sains Malaysia 11800 USM, Penang, Malaysia
Y. Abu Hasan
School of Mathematical Sciences, Universiti Sains Malaysia 11800 USM, Penang, Malaysia
Journal of Applied Sciences
Year: 2011 | Volume: 11 | Issue: 22 | Page No.: 3717-3724







ABSTRACT
In this study a mathematical model of two competing predators sharing one prey in homogeneous environment, with Holling type-I functional responses, was introduced. The model was formed by considering logistic law for both prey and predator where prey abundance was used to determine carrying capacity of predators. Stability of the equilibrium points of the system was then studied. The conditions for the persistency which were interpreted as efficiency of search and capture, were then obtained. Numerical simulations showed that the coexistence and extinction of the predators depend on how close the efficiency of searching and capture of the two predators.
PDF Abstract XML References Citation
Received: July 18, 2011;
Accepted: November 23, 2011;
Published: December 29, 2011
How to cite this article
Jawdat Alebraheem and Y. Abu Hasan, 2011. The Effects of Capture Efficiency on the Coexistence of a Predator in a Two Predators-One Prey Model. Journal of Applied Sciences, 11: 3717-3724.
DOI: 10.3923/jas.2011.3717.3724
URL: https://scialert.net/abstract/?doi=jas.2011.3717.3724
DOI: 10.3923/jas.2011.3717.3724
URL: https://scialert.net/abstract/?doi=jas.2011.3717.3724
INTRODUCTION
The study of interactions between species is a well researched field in ecology. In the case of one species (the predator) feeding on another species (the prey), a simple model was developed to understand the interaction. The model can be written as a coupled system of two first order non-linear differential equations (Giordano et al., 2003). Non linear differential equations are used in many fields (Taiwo and Abubakar, 2011; Baghli and Benyettou, 2006; Prakash and Karunanithi, 2009). The standard basic predator-prey model is the Lotka-Volterra system of equations (Rockwood, 2006). The major part of existing theory on predator-prey interactions is built on this basic model.
Extensions of the basic model were developed to describe particular scenarios and these involve the surroundings and the nature of the species involved. The resulting system of equations describing the predator-prey equations has been widely studied by many researchers (Naji and Balasim, 2007). However, there is still a need to improve the present models. In nearly all of these models, the main question that needs to be answered is whether the species can coexist.
Relative to the size of the prey, the size of the predators might involve smaller rate of change and this can result in competition amongst the predators (Mallah et al., 2001; Akhtar and Khaliq, 2003). This scene can be extended by considering the presence of another predator species preying on the same prey. Models involving more than two species have been proposed to describe some ecological phenomena, with very complex dynamical behaviors exhibited (Naji and Balasim, 2007; Lv and Zhao, 2008; Yu and Zhao, 2009; Upadhyay and Naji, 2009).
In this study, the dynamical relations of two predator species predating on a prey species were investigated. Various research approaches were undertaken to analyze this particular species configurations (Dubey and Upadhyay, 2004; Gakkhar et al., 2007). The approach here was slightly taken different from others. From the persistency conditions of the predators, the search and capture efficiency of the predators was defined. The developed model and the methods of analysis were able to answer some questions such as how the coexistence and extinction of the predators depend on the efficiency of the search and the capture. Several numerical simulations were carried out in the case of a non periodic solution.
MATHEMATICAL MODEL
According to Rockwood (2006), the diversity of organisms and the difference in the environments have led to different models of population growth. He added that the search for one model or one set of models for all population in different environments is pointless. With this in mind, a new model with the growth rates of the prey and two predators is described by the logistic law, with the carrying capacity of the predators depending on the available amount of prey is proposed. By using the Holling type-I functional response to describe the predation of the two predators y and z on prey x, the model can be written as:
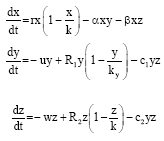 | (1) |
The intrinsic growth rate of prey is r.α and β measure the efficiency of search and capture of predators, z respectively. In the absence of prey x, constants u and w are the death rates of predators y, z respectively. R1 = e1αx, R2 = e1βx; R1 and R2 represent numerical responses of the predators y, z, respectively which describe change in the population of predators through prey consumption. e1 and e2 represent efficiency of converting consumed prey into predator births. The carrying capacities kx = a1x, kz = a2z are proportional to the available amount of prey, as was first proposed by Leslie (Gazi and Bandyopadhyay, 2008). c1 and c2 measure intraspecific competition between the predators.
The system of Eq. 1 can be written in a non-dimensional form. This can be done in many ways but it is better that the choice of variables relate to some key parameters. This makes the analysis less complicated as the number of parameters is reduced from 12 to 8. Using the following transformations of variables and parameters:
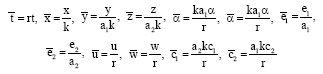 |
we have:
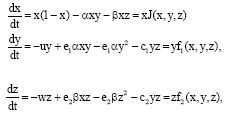 | (2) |
Equations 2 are of Kolgomorov type.
Theorem 1: The solution of the system (2) for t≥0 in ![]() is bounded.
is bounded.
Proof: The first equation of the system (2) that represents the prey equation is bounded through:
| (3) |
The solution of the equation (3) is:
where:
is the constant of integration. Hence x(t)+y≤1, ∀t≥0. Next, we prove thatx(t)+y(t)+z(t)≤L, ∀t≥0. Let D(t) = x(t)+y(t)+z(t). The time derivative of the function D:
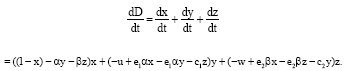 | (4) |
However, the solutions initiating remain in nonnegative quadrant in R3 and all the parameters are positive; it can be assumed the following:
| (5) |
It can be concluded that:
| (6) |
By substituting in Eq. 5 become as follows:
| (7) |
| (8) |
Equation 8 can be written as follows:
| (9) |
Since x(t)≤1, then:
| (10) |
But:
| (11) |
And:
| (12) |
Positive values are taken in (11) and (12).
So (10) becomes:
| (13) |
Where:
| (14) |
Consequently Z(t)≤L+σe-1, σ is a constant of integration. As t→ ∞ we have D(t)≤L.
EQUILIBRIUM POINTS AND STABILITY ANALYSIS
Equilibrium points: It is observed that the system (2) has five nonnegative equilibrium points. E0 = (0,0,0) and E1 = (0,0,0) are obvious (i.e. they exist without conditions on parameters):
and
exist and are positive if the following conditions hold:
| (15) |
| (16) |
The fifth positive equilibrium point ![]() will not be shown as it contains many parameters and hence will be very long.
will not be shown as it contains many parameters and hence will be very long.
Stability analysis: To study the local dynamical behavior of equilibrium points, the variational matrices of each equilibrium points are computed. From these matrices and using the Routh-Hurwitz criteria the local asymptotically stable are determined.
The variational matrix of E0 is given by:
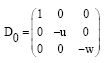 |
It is observed from D0 that the manifold is unstable along x-direction but stable along y-direction and along z-direction because the eigenvalue of x-direction is positive, while the eigenvalues of y-direction and z-direction is negative. Therefore, the first equilibrium point E0 is saddle point.
The variational matrix of E1 is given by:
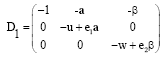 |
The equilibrium point E1 is locally asymptotically stable, provided the following conditions hold:
| (17) |
| (18) |
The variational matrix of E2 is given by:
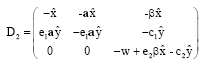 |
From D2 and by using the Routh-Hurwitz criteria it is observed that the equilibrium point E2 is locally asymptotically stable, provided the following conditions hold:
| (19) |
| (20) |
The equilibrium point E2 is stable in x-y plane if the condition (19) is satisfied. But the condition (15) must be satisfied so the condition (19) is always satisfied, while the stability of the E2 in the z direction (i.e. orthogonal direction to the x-y plane) depend on the condition (20).
The variational matrix of E3 is:
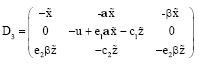 |
The equilibrium point E3 is locally asymptotically stable with the following condition:
| (21) |
| (22) |
The equilibrium point E3 is stable in x-z plane if the condition (21) is satisfied. On the other hand the condition (16) must be satisfied so the condition (21) is always satisfied, while the stability of the E3 in the y direction (i.e., orthogonal direction to the x-z plane) depends on the condition (22).
For the equilibrium points:
the variational matrix is:
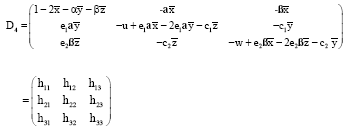 |
Where:
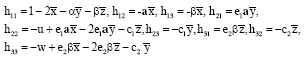 |
The characteristic equation of the variational matrix D4 is:
With:
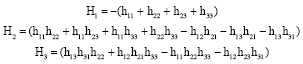 |
From Routh-Hurwitz criterion:
is locally asymptotically stable if the following conditions hold:
| (23) |
| (24) |
| (25) |
We thus proved the following theorem.
Theorem (2):
| • | The equilibrium point E0 = (0,0,0) is a saddle point with locally stable manifold in the y-z plane and with locally unstable manifold in the x direction |
| • | The positive equilibrium point E1 = (0,0,0) is locally asymptotically stable in the x-direction but it is locally asymptotically stable in x-z plane if it holds the conditions (17) and (18). The equilibrium point E1 is a saddle point if the conditions (17) and/or (18) are not satisfied |
| • | The equilibrium points: |
and:
| • | Are positive under the conditions (15) and (16) respectively. The equilibrium point E2 is locally asymptotically stable provided the conditions (19) and (20) hold, while the equilibrium point E3 is locally asymptotically stable provided the conditions (21) and (22) hold |
| • | The non-trivial positive equilibrium point |
| • | Exists; it is locally asymptotically stable provided the conditions (23) (24) and (25) hold |
Corollary: The equilibrium points E2 and E3 are unstable in z-direction (i.e. orthogonal direction to the x-y plane) and in y-direction (i.e. orthogonal direction to the x-z plane), respectively, if the condition (20) of E2 and the condition (22) of E3 are not satisfied (violated).
Theorem (3):
| • | The equilibrium point E2 is globally asymptotically stable inside the positive quadrant of x-y plane |
| • | The equilibrium point E3 is globally asymptotically stable inside the positive quadrant of x-z plane |
Proof: We prove (i) and in the same manner (ii) can be proved.
Let:
is a Dulac function, it is continuously differentiable in the positive quadrant of x-y plane, A = {(x,y)| x>0,y>0}.
Let:
Thus:
It is observed that Δ(GN1, GN2) is not identically zero and does not change sign in the positive quadrant of x-y plane A. So by Bendixson-Dulac criterion (Logan and Wolesensky, 2009), there is no periodic solution inside the positive quadrant of x-y plane. E2 is globally asymptotically stable inside the positive quadrant of x-y plane.
Persistence and extinction: Freedman and Waltman (1984) were studied system of equations of Kolgomorov type and derived abstract theorems that showed persistency once certain conditions were applied. They used a strong definition of persistency which is: A population x(t) is persistent if x(0) > 0 and lim inft → ∞ x(t)>0. The system is said to persist if each component of the system persists. The system (2) has no periodic solution in the respective planes as was shown in Theorem 3. Also, the boundedness of system (2) was proved in Theorem 1. According to Corollary 1, the orthogonal directions of E2 and E3 are unstable if the conditions (20) and (22) are not satisfied.
We next show that the conditions of Freedman and Waltman (1984) are satisfied. We use y1 = y and y2 = z to simplify the notations.
(C1) x is a prey population and y, z are competing predators, living exclusively on the prey, i.e:
(C2) In the absence of predators, the prey species x grows to carrying capacity, i.e.:
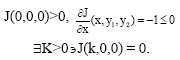 |
Here k = 1.
(C3) There are no equilibrium points on the y or z coordinate axes and no equilibrium point in y-z plane.
(C4) The predator y and the predator z can survive on the prey, This means that there exist points:
and:
such that:
and:
and ![]() .
.
Freedman and Waltman (1984) then showed that if the above conditions hold, if there is no limit cycle and if:
| (26) |
then system (2) persists.
Inequalities (26) implies that:
| (27) |
| (28) |
Gakkhar et al. (2007) has mentioned the conditions that represents the necessary conditions to include the following:
| (29) |
In the case of satisfying the conditions (27) and (28), then system (2) persists. However, in the case condition (27) was satisfied but condition (18) was not satisfied, then the first predator y survive, while the second predator z becomes extinct and vice versa.
Numerical simulations: Different values of the parameters α and β in studying the dynamical behavior of the system numerically were considered. The parameters α and β are important parameters because they are contained in the functional and numerical responses which formed the main component of prey predator models (Rockwood, 2006). In addition, they are part of the intraspecific competition coefficients in our model. The functional response plays an important role in interactions between prey and predator (Poggiale, 1998). α and β measure the efficiency of search and capture of the predators. Two different cases were considered which showed the coexistence and extinction of the predators. The other parameters were fixed in both cases. However, the numerical simulations focused on showing the coexistence or extinction of one of the predators. The values of parameters were chosen to satisfy the stability conditions of equilibrium points E2 and E3 in x-y, x-z planes, respectively which imply non periodic solution (Theorem 3). The other parameters were fixed as follows:
Two different sets of numerical experiments were carried out. In the first case, the value of β was fixed at 1.33 and the value of α varies. It is observed in Fig. 1 that when the values of α and β were near to each other, the three species stably coexist.
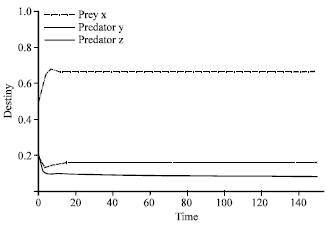 | |
| Fig. 1: | Time series of dynamical behavior of the system (2) at α = 1.37. Figure 1 described the coexistence of three species (two predators one prey system) when α which is the value of efficiency of search and capture of predator y was near of β that is efficiency of search and capture of predator z, where the value of α was 1.37. The dot dashed line represents prey x, the dashed line indicates predator y, while the connected thick line represent predator z, this system of lines is applied in all figures |
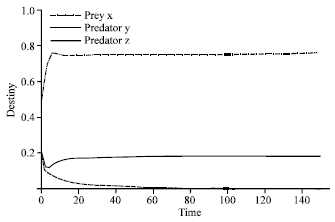 | |
| Fig. 2: | Time series of dynamical behavior of the system (2) at α = 0.9. Fig. 2 showed extinction of predator y and surviving predator z when the value of efficiency of search and capture of predator y (α) was decreased. Predator z is represented by the connected thick line, predator y is indicated by the dashed line, while prey x is signified by the dot dashed line |
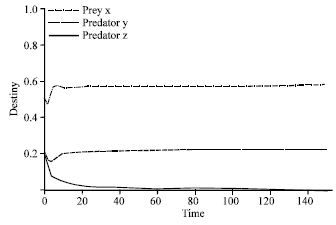 | |
| Fig. 3: | Time series of dynamical behavior of the system (2) at α = 1.9. Figure 3, extinction of predator z and surviving predator y was shown when increasing the value of efficiency of search and capture of predator y (α) to become 1.9. The dashed line indicates predator y, predator z is represented by the connected thick line, while the dot dashed line represents prey x |
However, if the value of α was decreased (α = 0.9), predator y became extinct, while predator z survived, as is shown in Fig. 2. On the other hand, if α was increased (α = 1.9), the predator z tended to extinction (Fig. 3) and predator y survived. This showed that the survival of each predator depended on the efficiency of the search and capture.
In the second case, the same values of parameters were used but the value of α was fixed at 1.1 and giving different values to β.
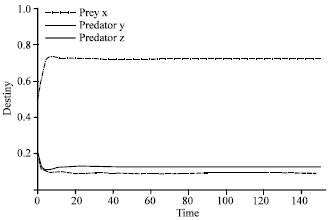 | |
| Fig. 4: | Time series of dynamical behavior of the system (2) at β = 1.3. In Fig. 4, the coexistence of three species (two predators one prey system) is shown when β that represents the value of efficiency of search and capture of predator z is near of value β which is efficiency of search and capture of predator y, the value of β is 1.3. The dashed line represents predator y, the connected thick line represent predator z, while the dot dashed line represents prey x |
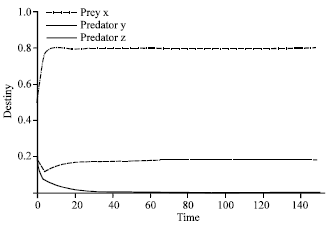 | |
| Fig. 5: | Time series of dynamical behavior of the system (2) at β = 0.9. Figure 5 illustrates extinction of predator z and surviving predator y when decreasing the value of efficiency of search and capture of predator z (β) to become 0.9. The dashed line indicates of predator y, predator z is represented by the connected thick line, while the dot dashed line represents prey x |
Corresponding results for β are shown in Fig. 4, Fig. 5 and Fig. 6. when the values of α and β were almost near to each other where three species may coexist at these values as it is shown in Fig. 4. But in case the value of β(β = 0.9) was decreased, predator z became extinct, while predator y survived, as is clear in Fig. 5. Otherwise if β was increased (β = 1.8), the predator y tends to extinction and predator z survives, this is explained in Fig. 6.
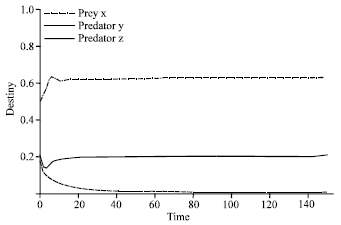 | |
| Fig. 6: | Time series of dynamical behavior of the system (3) at β = 1.8. Figure 6 shows surviving predator z and extinction of predator y when increasing the value of β to become 1.8. The connected thick line represents predator z, the dashed line indicates of predator y, while the dot dashed line represents prey x |
It was observed that the numerical simulations correspond with theoretical analysis when the conditions in section 5 were applied.
Predators and prey can influence one another's evolution. Traits that enhance a predator's ability to find and capture a prey will be selected in the predator. Traits that enhance a prey's ability to avoid being captured and eaten by a predator will be selected in the prey. The “goals” of these traits are not compatible and it is the interaction of these selective pressures that influences the dynamics of the predator and prey populations (El-Messoussi et al., 2007; Younas et al., 2004; Islam et al., 2004). Predicting the outcome of species interactions is also of interest to biologists trying to understand how communities are structured and sustained. One aspect that may affect the efficiency of searching and capture by predators is the existing environmental effects interfere with their foraging activities (Myers et al., 2007; Smee, 2010). Recent research has shown that the movement properties of foraging animals may have important implications for their success in locating prey (Scharf et al., 2006).
CONCLUSIONS
In this study a continuous time mathematical model of interactions of two competing predators sharing one prey was introduced. Holling type-I functional responses have been used. The conditions of existence of equilibrium points and their stability of equilibrium points of the model were obtained. Theoretical analysis on persistence of the system and the extinction of one of the predators was presented.
Numerical simulations showed that if the efficiency of searching and capture of both predators was roughly the same, the three species can co-exist. However, if the efficiency of one of the predator was less than the other, this leaded to the extinction of the later predator.
ACKNOWLEDGMENTS
This research is supported by a grant from the School of Mathematical Sciences, Universiti Sains Malaysia.
REFERENCES
- Akhtar, I.H. and A. Khaliq, 2003. Impact of plant phenology and coccinellid predators on the population dynamic of rose aphid Macrosiphum rosaeiformis das (Aphididae: Homoptera) on rose. Asian J. Plant Sci., 2: 119-122.
CrossRefDirect Link - Baghli, H. and M. Benyettou, 2006. Modeling and simulation of thermal transfer in batch reactor. J. Applied Sci., 6: 1327-1331.
CrossRefDirect Link - Dubey, B. and R.K. Upadhyay, 2004. Persistence and extinction of one-prey and two-predators system. Nonlinear Anal. Model. Control, 9: 307-329.
Direct Link - El-Messoussi, S., H. Hafid, A. Lahrouni and M. Afif, 2007. Simulation of temperature effect on the population dynamic of the Mediterranean fruit fly Ceratitis capitata (diptera: tephritidae). J. Agron., 6: 374-377.
CrossRefDirect Link - Gakkhar, S., B. Singh and R.K. Naji, 2007. Dynamical behavior of two predators competing over a single prey. BioSystems, 90: 808-817.
CrossRef - Gazi, N.H. and M. Bandyopadhyay, 2008. Effect of time delay on a harvested predator-prey model. J. Applied Math. Comput., 26: 263-280.
CrossRef - Islam, M.S., M.U. Ahmad, A.H.M.M. Haque and M.E.H. Sarker, 2004. Population dynamics of Hirschmanniella oryzae in the rice root of farmer fields as affected by edaphic factors. Pak. J. Biol. Sci., 7: 2002-2004.
CrossRefDirect Link - Lv, S. and M. Zhao, 2008. The dynamic complexity of a three species food chain model. Chaos, Solitons Fractals, 37: 1469-1480.
CrossRef - Mallah, G.H., A.K. Korejo, A.R. Soomro and A.W. Soomro, 2001. Population dynamics of predatory insects and biological control of cotton pests in Pakistan. J. Biol. Sci., 1: 245-248.
CrossRefDirect Link - Myers, R.A., J.K. Baum, T.D. Shepherd, S.P. Powers and C.H. Peterson, 2007. Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science, 315: 1846-1850.
CrossRefDirect Link - Naji, R.K. and A.T. Balasim, 2007. Dynamical behavior of a three species food chain model with Beddington-DeAngelis functional response. Chaos, Solitons Fractals, 32: 1853-1866.
CrossRef - Prakash, N. and T. Karunanithi, 2009. Advances in modeling and simulation of biomass pyrolysis. Asian J. Scientific Res., 2: 1-27.
CrossRefDirect Link - Poggiale, J.C., 1998. Predator-prey models in heterogeneous environment: Emergence of functional response. Mathl. Comput. Model., 27: 63-71.
CrossRef - Scharf, I., E. Nulman, O. Ovadia and A. Bouskila, 2006. Efficiency evaluation of two competing foraging modes under different conditions. Am. Nat., 168: 350-357.
PubMed - Smee, D., 2010. Environmental context influences the outcomes of predator-prey interactions and degree of top-down control. Nat. Educ. Knowledge, Vol. 1.
Direct Link - Taiwo, O.A. and A. Abubakar, 2011. An application method for the solution of second order non linear ordinary differential equations by chebyshev polynomials. Asian J. Applied Sci., 4: 255-262.
CrossRef - Upadhyay, R.K. and R.K. Naji, 2009. Dynamics of a three species food chain model with Crowley: Martin type functional response. Chaos, Solitons Fractals, 42: 1337-1346.
CrossRef - Younas, M., M. Naeem, A. Raqib and S. Masud, 2004. Population dynamics of cabbage butterfly (Pieris brassicae) and cabbage aphids (Brevicoryne brassicae) on five cultivars of cauliflower at Peshawar. Asian J. Plant Sci., 3: 391-393.
CrossRefDirect Link - Yu, H. and M. Zhao, 2009. Dynamic behavior of a three-species ecological system with the beddington-deangelis functional response. Proceedings of the International Workshop on Chaos-Fractals Theories and Applications, November 6-8, 2009, Shenyang, pp: 346-350.
CrossRef