
K. Zaleha
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, Malaysia
I. Hazwani
Department of Marine Science, Universiti Malaysia Terengganu, Malaysia
H. Siti Hamidah
Department of Marine Science, Universiti Malaysia Terengganu, Malaysia
B. Y. Kamaruzzaman
Kulliyyah Science of International Islamic University Malaysia, Kuantan, Pahang, Malaysia
K. C. A. Jalal
Kulliyyah Science of International Islamic University Malaysia, Kuantan, Pahang, Malaysia
Journal of Applied Sciences
Year: 2011 | Volume: 11 | Issue: 14 | Page No.: 2620-2626







ABSTRACT
Tachypleus gigas (Muller) the Southeast Asian horseshoe crab, is found to nest on sandy beaches in Malaysia, thus biological information about survival of this living fossil become crucial. Laboratory experiments were carried out to investigate the effect of salinity on duration to maturation and hatching success of their eggs. The effect on size, morphological changes and growth of the hatched larvae was also investigated. Eggs of T. gigas were collected from the newly nests made on breeding beach of Pekan, Pahang (Lat 3° 56.915’N; Long 103°21.933’ E) in Peninsular Malaysia. Triplicates of five different salinities (15, 20, 25, 30 and 35 ppt) were chosen for the experiments. A vernier caliper was used for size measurement under a stereo microscope. The morphological changes in prosomal width and length, body length, telson length, opisthosoma length and total length were recorded. Salinity range of between 25 and 35 ppt could be an optimal incubation condition. No egg hatched at salinity 15 and 20 ppt. Highest percentage of hatching was found at 30 ppt (91.11±5.57%). Hatching also occurred at 25 and 35 ppt with 65.56±1.73% and 74.4±2.31% success rate, respectively. The shortest molting time occurred at 15 ppt (25 days after hatching). Total size changes in larvae were also highest at 15 ppt but lowest at 35 ppt. There was no mortality in larvae maintained in all treatments. Early larval development would also affected by the salinity stress but its seemed tolerable.
PDF Abstract XML References Citation
Received: November 14, 2010;
Accepted: April 12, 2011;
Published: June 07, 2011
How to cite this article
K. Zaleha, I. Hazwani, H. Siti Hamidah, B. Y. Kamaruzzaman and K. C. A. Jalal, 2011. Effect of Salinity on the Egg Hatching and Early Larvae of Horseshoe Crab Tachypleus gigas (Muller, 1785) in Laboratory Culture. Journal of Applied Sciences, 11: 2620-2626.
DOI: 10.3923/jas.2011.2620.2626
URL: https://scialert.net/abstract/?doi=jas.2011.2620.2626
DOI: 10.3923/jas.2011.2620.2626
URL: https://scialert.net/abstract/?doi=jas.2011.2620.2626
INTRODUCTION
Horseshoe crab, a chelicerate arthropod, is an ancient animal survived from ancient time to date, thus called as living fossil. The extraordinary defense mechanism is due to the blood property in this animal. It has now becomes a great contribution to the world of medical and pharmaceutical science (John et al., 2010). Limulus polyphemus, a species endemic to Atlantic Ocean has been reported to be over-exploited and now strictly monitored and regulated due to its importance in the pharmaceutical industry worldwide. Only licensed biomedical companies could purchase large crabs, which are harvested by trawlers or by hand from spawning beaches (ASMFC, 2004).
Abundance of other horseshoe crab species is also reported to decrease tremendously (Tanacredi, 2001; Chen et al., 2004). The development for human need in Asia as well has extremely put on stress to horseshoe crab found in this region, thus giving the status of the three species found in Asia as under threat, the same as the Atlantic species (Morton and Blackmore, 2001). Tachypleus gigas (Muller) is listed under Data Deficient in the 2009’s IUCN together with Carcinoscorpius rotundicaula (Latreille) and Tachypleus tridentatus (Leach).
T. tridentatus, T. gigas and C. rotundicaula are found in Malaysia.They were reported under threat of unregulated harvesting for commercial purposes. T. tridentatus is reported only in Sabah water, East Malaysia. The other two species are commonly found in Peninsular Malaysia and could be distinguished physically by the body size and shape of tail (Kassim et al., 2008). T. gigas is bigger in size with triangular in cross section of tail while C. rotundicauda being smaller in size with cylindaric-shaped tail.
Horseshoe crab of Tachypleus is a marine species with the smaller juveniles prefer to be close to the shore than the older juveniles (Hu et al., 2009). Sediment texture indirectly influences horseshoe crab egg viability and development (Jackson et al., 2005) and Chiu and Morton (2003) suggested that the sediment with medium-sized sand and moderately sorted condition would promote hatching success and survival of horseshoe crab larvae on beach. Ehlinger and Tankersley (2003) was previously noted that hydration and agitation with sand may increase hatching rate. Apparently they have adopted a life strategy to survive on sandy beach that is exposed to fluctuation of temperature and salinity (Chiu and Morton, 2004). During spawning season in Pahang, Peninsular Malaysia, T. gigas was reported to build up to 22 nests on the breeding beach and nests could contained up to 407 individual eggs or larvae (Nur Erni et al., 2010). Although there was fluctuation in salinity on the breeding beach, there was no correlation found with the number of nest or eggs in their study area.
Report on the effect of salinity on the eggs, hatching success or larval development of horseshoe crab from tropical coast is scarce. The hatching success at beaches in Malaysia is not known to date. Furthermore, the conservation status of horseshoe crab in Malaysia as well as in many parts in Asia is still unknown while the populations continuely facing threats from potential loss of prime spawning habitat because of erosion and/or coastal development, and pollution (Botton, 2001). Pollution is reported as the main factor affecting the development of embryos and larvae of Tachypleus gigas and Carcinoscopius rotundicauda rather than salinity and temperature (Botton and Itow, 2009). Despite that, a study on the larval survival and development in T. gigas from India showed the important role of salinity (Chatterji et al., 2004). In view of the long term needs to conserve and manage this important species in Malaysia water, and the potential contribution of culture success of the larval and juvernile stage for conservation, effort to compile data on their ecology and biology should become priority as to fully understand their biological importance. In the present study, focus is made to investigate the effect of different salinity towards the eggs, their hatching success and early larval growth under laboratory condition. This information is vital for future work on conservation and beach rehabilitation program.
MATERIALS AND METHODS
Field sampling: Field sampling was carried out in day time during the full moon of August 2009 at the breeding beach of Pekan, Pahang (Latitude 3° 56.915’N; Longitude 103°21.933’ E) during low tide. Station for field sampling, Pekan, Pahang (Latitude 3° 56.915’N, Longitude 103°21.933’) was located in the East Coast of the Malay Peninsula, facing the South China Sea. The field sampling was done for two days, 5-7th of August 2009 at about noon which was coincide with the spring tide. It was during the full moon of lunar cycle where high number of horseshoe crabs could be expected to land for breeding on the beach. Field survey was started about 2hours before the time of highest tide. Adults horseshoe crabs were observed to come out to the beach following the tide.
Spawning pairs of horseshoe crab T. gigas were monitored and the nests left by the pairs were escavated carefully. During the field work, observation was made from far and the spawned females were left undisturbed or captured, thus no data on female size was collected. Horseshoe crab eggs were collected by hand. About 1500 of horseshoe crab eggs were obtained for this study. At the sampling site, the eggs were taken with sand from their nest as to ensure the sprayed sperms from male fully fertilised the eggs. The physical parameter namely the Dissolved Oxygen (DO), pH, salinity and temperature at the sampling site were measured in situ using a YSI 556 multi-parameter probe. Color of eggs was observed and recorded and eggs counting were done at the sampling site.
Sieve of 500 microns was used to discard the sand and other particles. The fertilized eggs are olive green in color with thick elastic chorion. Horseshoe crab eggs were then transferred into plastic bag containing filtered natural seawater (30 ppt). As no technical aeration was provided, the plastic bag was opened to let dissolved of oxygen from air periodically during their transportation to laboratory.
Laboratory experiment 1 and 2 was then carried out in the marine hatchery of Institute of Tropical Aquaculture, Universiti Malaysia Terengganu. Upon arrival, all eggs were transferred into containers with filtered natural seawater of 30 ppt and mild aeration. The eggs were maintained for a week before experiment in ambient temperature (26-29°C). All experiments were carried out within two months.
Experiment 1
Effect of salinity on egg development and hatching success: Artificial Sea Water (ASW) was prepared to get the salinity of 15, 20, 25, 30 and 35 ppt in triplicates glass beaker container of 500 mL. Salinity was measured using a salinity refractometer (model RHS10 ACT) with offered feature of Automatic Temperature Compensation. The refractometer was first calibrated by placing 2 to 3 drops of distilled water on the meter, look through the eyepiece and turn the calibration screw until the scale reads 0. To check the salinity of the prepared beakers, 2 to 3 drops of water from each beaker were placed on the meter before reading was made through the eyepiece. Soft aeration was given to each container. A total of 30 fertilized eggs were carefully placed in each beaker. Each beaker was then covered with parafilm as to reduce dessication. The experiment was carried out in room temperature (27-29°C) under natural light with more or less of natural photoperiod. No additional lighting was given.
Physical parameters were measured every day using YSI Meter Model 550A for temperature and dissolved oxygen, a portable pH meter (Thermo Russel RL060p) for pH and a refractometer (model RHS10 ACT) for salinity. ASW was changed every other day and samples were checked daily for the presence of newly hatched larvae. A vernier caliper was used for size measurement. The egg development was observed under a stereo microscope and hatched eggs were recorded.
Experiment 2
Effect of salinity on early larval growth: Eggs collected from field sampling were maintained at 30 ppt in laboratory condition. Culture condition for this experiment was as the same as in experiment 1. A total of 30 newly hatched larvae were put into each 500 mL experimental glass beaker of 15, 20, 25, 30 and 35 ppt and softly aerated. Larvae were fed with a planktonic copepod Apocyclops sp. at a rate of 26 copepod/day/larvae. Supply of copepods was obtained from the established culture in the same laboratory.
The physical parameters of culture in all beakers were measured daily as in Experiment 1 for 40 days. To count the survival rate, any mortality was recorded everyday. The larvae were considered died when they were inactive (no leg or book-gill movement) and did not response to mechanical stimulation. The morphological changes were observed and described as change of size in prosomal width and length, body length, telson length, opisthosoma length and total length.
Data collected was analysed statistically to determine the effect of different salinity on development, duration of eggs to hatch and hatching success using SYSTAT 11.5, SPSS. One-way Analysis of Variance (ANOVA) was used to determine the significant difference of time needed for hatching and the larvae to molt into first instar. It is also used to test if the size changes of body parts were significantly different at different salinities.
RESULTS
Environmental parameters: Environmental parameters measured in situ during the field sampling is summarizes in Table 1. The values indicate the sampling area was at a normal condition of tropical water. Dissolved oxygen showed comparatively low concentration while temperature indicated as warm weather condition during the samplings.
In the laboratory, culture condition was maintained at almost the same condition found in their nest except for temperature (Table 2).
| Table 1: | Salinity, Dissolved Oxygen, pH, temperature (°C) and conductivity (mS cm-1) of overlying water at horseshoe crab nest at Pekan Pahang |
| Table 2: | Mean value of DO (mg L-1), pH and temperature (°C) maintained in different salinity |
 | |
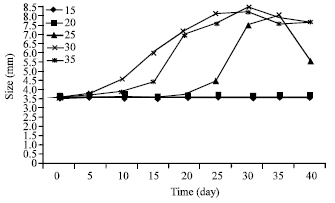 | |
| Fig. 1: | Effect of salinity (ppt) on size of T. gigas’s egg maintained at laboratory condition |
Temperature in the laboratory was maintained at between 26.7±0.2 and 27.2±0.1°C, lower than at field.
Effect of different salinity on eggs size: Initially, at all salinities, the increment in size was slow but at day-10 eggs maintained in 30ppt increase their size drastically (Fig. 1). On the day-1, eggs were 3.58±0.06 mm in diameter. On the day-25, the egg size was bigger at salinity 30 ppt before reduced on the day-35. Maximum mean size attained by the eggs was recorded at salinity 30 ppt (6.2 mm). There was no size increase at salinity 15 and 20 ppt.
Effect of different salinity on hatching success: The analysis of variance showed a significant different of day to hatch (p<0.05) between salinity treatment. No eggs hatched at salinity 15 and 20 ppt.The shortest duration for the eggs to hatch was when maintained at 30 ppt with the mean duration of about 33 days (Table 3). Number of individual to firstly hatch was also highest at 30ppt. At this salinity, the earliest times for the eggs to hatch was on the day-26 with 8 individuals (27%). Higher number of hatched eggs was obtained on the day-27 with 11 individuals (37%) (Fig. 2). At salinity 35 ppt, horseshoe crab eggs started to hatch on the day-28 with 7 individuals (23%) and at salinity 25 ppt it hatched on the day-30 with 9 individuals (30%).
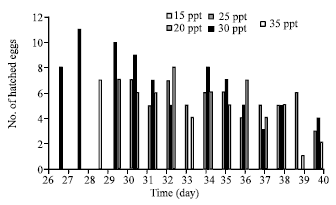 | |
| Fig. 2: | Effect of salinity (ppt) on number of hatch per day and day to hatch of T. gigas’s egg maintained in laboratory |
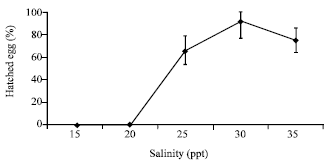 | |
| Fig. 3: | Effect of salinity (ppt) on hatching success of T. gigas’s egg maintained in laboratory condition |
| Table 3: | Mean number of days for eggs to hatch at different salinities |
 | |
Effect of salinity on total percentage of hatching success is summarizes in Fig. 3. Highest percentage was found at 30 ppt (91.11±5.57%) while at 35 and 25 ppt the hatching success was at 74.4±2.31% and 65.56±1.73%, respectively. No hatching was observed in both the 15 and 20 ppt and eggs had already covered by fungi. After day-35, most of horseshoe crab eggs in other salinities became rotten eggs and the color turned red or black.
Effect of salinity on growth of larvae: Trilobites hatched from the eggs was monitored for their growth within 40 days and the result on the changes in different morphometric data is summarizes in Fig. 4. Salinities of 15 to 25 ppt showed stronger effect on the size changes if compared to at 30 and 35 ppt.
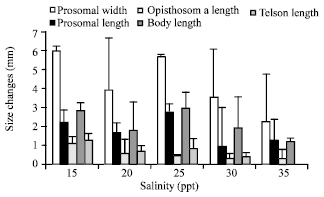 | |
| Fig. 4: | Size changes as shown by different morphometric data of trilobites of T. gigas maintained in different salinities |
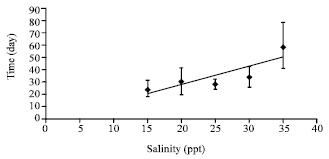 | |
| Fig. 5: | Effect of salinity (ppt) on molting time of trilobites of T. gigas maintained in laboratory condition |
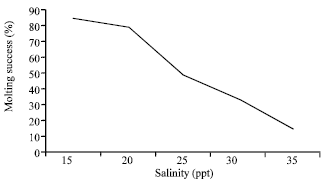 | |
| Fig. 6: | Effect of salinity (ppt) on molting success (%) of trilobites of T. gigas maintained in laboratory condition |
Major size change for every treatment was shown by the prosomal width followed by the body length while the least was by telson length.
The time needed by the trilobites to molt into instars increases significantly (p<0.05) with the increase of salinity. Due to their fast increase in size, the larvae at 15 ppt took the shortest time to molt into first instar which was only 25 days after hatching (Fig. 5). At 20 and 25 ppt, it took 30 and 28 days, respectively to molt while at 30 ppt, molting occurred on day-34. No molting larvae were observed within 40 days experiment at 35 ppt.
Percentage of molting success in different salinity is shown in Fig. 6. Obviously the molting success decreases to less than 50% if cultures were maintained at higher salinity between 30 and 35 ppt. In contrast, higher molting percentage was found when trilobites were maintained at salinity 15 and 25 ppt.
DISCUSSION
Effect of different salinity on egg size and hatching: Salinity fluctuations are common in beaches and it is known to influence survival and growth of marine invertebrates including horseshoe crab (Botton et al., 2006). Its influences many physiological functions and important in regulating the distribution of estuarine and marine organisms (Laughlin and French, 1989; Goncalves et al., 1995). Ehlinger and Tankersley (2004) found that embryo in the eggs of L. polyphemus could develop into larval stage and successfully hatch in salinities between 10 and 70 ppt. In contrast, eggs of T. gigas failed to increase their size or hatch at salinity of 15 and 20 ppt in the present study which could be related to the small tolerant limit and salinity fluctuation in this tropical species.
The change in inner fluid osmolality in response to outer medium osmalality had been reported in crustacean such as homarid lobsters (Charmantier et al., 2001). This possibly true also for horseshoe crab due to the nature of horseshoe crab’s embryonic membrane which was reported as very permeable to water, chloride and sodium ions (Laughlin, 1981). Hannan and Evans (1973) already proved the ability of Limulus to change it permeability when maintained at low salinity medium, a strategy of invertebrate to regulate osmotic pressure in relation to the external environment (Prusch, 1983). Ehlinger and Tankersley (2004) related the hyperosmotic condition of internal fluid during development to the osmotic shock condition as a result of certain factor such as dessiccation. Nevertheless, there was no shocking condition applied to the eggs in the present study and size increment was only occurred at salinities 25, 30 and 35 ppt. The size increase must be related to the increase in the embryo development which occur at it salinity tolerence limit (25-35 ppt) and salinity 15 and 20 ppt could be beyond the tolerant limit for T. gigas’s eggs in this study. This is shown by the highest percent of hatching found at 30 ppt while less number of hatching occurred at 25 and 35 ppt. This results were in agreement with Kosaku et al. (2000) where salinity of higher than 19 ppt was reported as suitable condition for T. tridentatus eggs to hatch, thus support the idea of a different salinity tolerant limit for different crab species.
Embryo of L. polymephus hatched into trilobites in 28 days before entered water during high tide (Penn and Brockmann, 1994). Trilobite larvae of T. gigas in India was reported to hatch out 40-45 days after fertilization (Chatterji et al., 2004). Nevertheless, by maintaining salinity at 30ppt at constant temperature in the present study, hatching occurred as early as 26th day after fertilization. This results were contradicting with those previous report most likely due to the different environmental setting for the study. Constant temperature maintained in the laboratory had reduced the possible thermal effect as usually experienced by the eggs at the beaches (Chiu and Morton, 2004). On the beach, the timing of hatching may be influenced by a counteracting influence of low salinity on embryonic development rate and hatching, particularly with the effect of freshwater run-off. At low salinities, embryos must have developed more slowly, but later the enhanced water content of the eggs might have accelerated the hatching process (Saigusa, 1996).
Effects of salinity on larval growth and molting: Morphometric data such as prosomal width and length is usually the practical way to measure growth in larval and juveniles of horseshoe crab (Lee and Morton, 2005). It will give important data such as the age of the horseshoe crab (Sekiguchi et al., 1988). Embryo and larval growth in L. polymephus showed that salinity influenced the rate of crab development, although only the extreme condition has an effect on survival (Ehlinger and Tankersley, 2004). They also indicated that the time from hatching to molting to the first juvenile instar decreased significantly with increasing salinity. Sekiguchi et al. (1988) found that growth of trilobite larvae and juveniles was slightly delayed at 35 ppt. These reports were in agreement with the present study where lower salinity of 15 and 25 ppt had initiate the increase in size of larvae particularly the prosomal width and body length. Giménez and Anger (2003) also noted on the same effect of incubation in low salinity to enhance the development of esturine crab larvae of Chasmagnathus granulata. They related the intraspecific variability in maternal energy investment into offspring production may play a significant role for the chances of larval survival and development.
The earliest molting period which was found at 15 and 30 ppt in the present study contradicting the report of Chatterji et al. (2004). In their study, at 40 ppt the growth rate was faster and moulting occurred on the 35th day of the experimental period, whereas moulting in other salinities (10, 20, 30 ppt) was recorded on the 42nd day. A recent report on the respiratory metabolism of trilobites of T. gigas indicating insignificant influence of salinity (10-40 ppt) on the oxygen consumption by the larvae (Suniza et al., 2011). This might indicate the role of other environmental parameters such as oxygen and temperature which affect larval growth. On the other hand, diet could be a factor that promote early molting. Jegla and Costlow (1970) maintained newly hatched trilobite larval culture of Limulus in seawater at constant temperature of 25°C and fed with newly hatched Artemia salina. Chatterji et al. (2004) did not offer any diet to their larval culture. There was no report on the effect of early diet to trilobite larvae in culture experiment but the copepod given during the experiment could be a contributor to the fast grow of the larvae. Copepods is known to have higher nutrient content than Artemia (Nanton and Castell, 1998; Carli et al., 1995). Further study is needed to confirm this effect. It was reported that in natural environment, the larvae usually bury themselves in the sand during high tide and emerge from it and begin to feed on the larger elements such as insect larvae or polychaete larvae in the surface sediments during low tide (Chiu and Morton, 2004).
CONCLUSION
The data collected in the present study adds new information on the incubation period of eggs of T.gigas in laboratory culture. Maintaining the eggs in constant salinity of its tolerence limit between 25 and 35 ppt particularly at 30 ppt could induce early hatching, as early as 26 days after fertilization. Trilobites reconditioned at low salinity of 15 ppt after hatching could increase their tendency to molt early, while higher salinities such as 35 ppt could delay molting process.
ACKNOWLEDGMENT
The authors are indebted to the Ministry of Agriculture and Agrobased Industry Malaysia (MOA) for the financial support through e-science fund granted to one of the authors.
REFERENCES
- Carli, A., G.L. Mariottini and L. Pane, 1995. Influence of nutrition on fecundity and survival in Tigriopus fulvus fischer (Copepoda: Harpacticoida). Aquaculture, 134: 113-119.
CrossRefDirect Link - Chatterji, A., S. Kotnala and R. Mathew, 2004. Effect of salinity on larval growth of horseshoe crab, Tachypleus gigas (Muller). Curr. Sci., 87: 248-249.
Direct Link - Chen, C.P., H.Y. Yeh and P.F. Lin, 2004. Conservation of the horseshoe crab at Kinmen Taiwan: Strategies and practices. Biodiversity Conservat., 13: 1889-1904.
Direct Link - Chiu, H.M.C. and B. Morton, 2003. The sediment and hydrographic characteristics of three horseshoe crab nursery beaches in Hong Kong. J. Ocean Univ. Qingdao, 2: 35-43.
Direct Link - Chiu, H.M.C. and B. Morton, 2004. The behaviour of juvenile horseshoe crabs, Tachypleus tridentatus (Xiphosura), on a nursery beach at Shui Hau Wan, Hong Kong. Hydrobiologia, 523: 29-35.
CrossRef - Ehlinger, G.S. and R.A. Tankersley, 2003. Larval hatching in the horseshoe crab, Limulus polyphemus: Facilitation by environmental cues. J. Exp. Mar. Biol. Ecol., 292: 199-212.
CrossRef - Ehlinger, G.S. and R.A.Tankersley, 2004. Survival and development of horseshoe crab (Limulus polyphemus) embryos and larvae in hypersaline conditions. Biol. Bull., 206: 87-94.
Direct Link - Goncalves, F., R. Ribeiro and A.M.V.M. Soares, 1995. Laboratory study of effects of temperature and salinity on survival and larval development of a population of Rhithropanopeus harrisii from the Mondego River estuary, Portugal. Mar. Biol., 121: 639-645.
CrossRef - Hannan, J.V. and D.H. Evans, 1973. Water permeability in some euryhaline decapods and Limulus polyphemus. Comp. Biochem. Physiol. A: Physiol., 44: 1199-1213.
CrossRef - Hu, M., Y. Wang, Y. Chen, S.G. Cheung, P.K.S. Shin and Q. Li, 2009. Summer distribution and abundance of juvenile chinese horseshoe crabs Tachypleus tridentatus along an intertidal zone in southern China. Aquat. Biol., 7: 107-112.
CrossRef - Jegla, T.C. and J.D. Costlow, 1982. Temperature and Salinity Effects on Developmental and Early Post-hatch Stages of Limulus. In: Physiology and Biology of Horseshoe Crabs: Studies on Normal and Environmentally Stressed Animals, Bonaventura, J., C. Bonaventura and S. Tesh (Eds.). Alan R. Liss Inc., New York, USA., pp: 103-113.
- John, B.A., K.C.A. Jalal, Y.B. Kamaruzzaman and K. Zaleha, 2010. Mechanism in the clot formation of horseshoe crab blood during bacterial endotoxin invasion. J. Applied Sci., 10: 1930-1936.
CrossRefDirect Link - Laughlin, R.B., 1981. Sodium, chloride and water exchange in selected developmental stages of the horseshoe crab Limulus polyphemus (Linnaeus). J. Exp. Mar. Biol. Ecol., 52: 135-146.
CrossRef - Laughlin, R.B. and W. French, 1989. Differences in responses to factorial combinations of temperature and salinity by zoeae from two geographically isolated populations of the mud crab Rhithropanopeus harrisii. Mar. Biol., 102: 387-395.
CrossRef - Lee, C.N. and B. Morton, 2005. Experimentally derived estimates of growth by juvenile Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura) from nursery beaches in Hong Kong. J. Exp. Mar. Biol. Ecol., 318: 39-49.
CrossRef - Nanton, D.A. and J.D. Castell, 1998. The effects of dietary fatty acids on the fatty acid composition of harpacticoid copepods, Tisbe sp., for use as a live food for marine fish larvae. Aquacult., 163: 251-261.
CrossRef - Nur Erni, A.H.N., K. Kamaliah, M.O. Luthfi, A. Chatterji, B.Y. Kamaruzzaman and K. Zaleha, 2010. Variation in distribution of horseshoe crab nest along pahang coastal area. Proceedings of the University Malaysia Terengganu International Annual Symposium Science and Management 2010, May 5-8, Kuala Terengganu, pp: 540-544.
- Penn, D. and H.J. Brockmann, 1994. Nest-site selection in the horseshoe crab, Limulus polyphemus. Biol. Bull., 187: 373-384.
Direct Link - Prusch, R.D., 1983. Evolution of invertebrate homeostasis: Osmotic and ionic regulation. Comp. Biochem. Physiol. A: Physiol., 76: 753-761.
CrossRef - Sekiguchi, K., H. Seshimo and H. Sugita, 1988. Post-embryonis development of the horseshoe crab. Biol. Bull., 174: 337-345.
Direct Link - Suniza, A.M., K. Zaleha and A. Chatterji, 2011. Effects of different environmental parameters on the respiratory metabolism of the larvae of Malaysian Horseshoe crab, Tachypleus gigas (Muller). Pertanika J. Sci. Technol., 19: 1-9.
Direct Link - Kosaku, M., S. Satoko, N. Shigetomo and H. Asuka, 2000. Ecology of hatched larvae of the horseshoe crab Tachypleus tridentatus (Leach) in relation to the physical environment. Jap. J. Benthol., 55: 15-24.
Direct Link - Gimenez , L. and K. Anger, 2003. Larval performance in an estuarine crab, Chasmagnathus granulata, is a consequence of both larval and embryonic experience. Mar. Ecol. Prog. Ser., 249: 251-264.
Direct Link - Botton, M.L.M. Pogorzelska, L. Smoral, A. Shehata and M.G. Hamilton, 2006. Thermal biology of horseshoe crab embryos and larvae: A role for heat shock proteins. J. Exp. Mar. Biol. Ecol., 336: 65-73.
CrossRef