
P. Abdeshahian
School of Bioscience and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
N. Samat
Livestock Research Center, Malaysian Agriculture Research and Development Institute, Mardi Headquarters, P.O. Box 12301, GPO, 50774 Kuala Lumpur, Malaysia
W.M. Wan Yusoff
School of Bioscience and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 5 | Page No.: 419-424







ABSTRACT
Aspergillus niger FTCC 5003 was cultivated on palm kernel cake as a substrate to produce β-xylosidase in solid substrate fermentation. The concurrent effects of three independent variables, including incubation temperature, initial moisture content of substrate and airflow rate on the production of β-xylosidase were studied by response surface methodology. A number of 18 trials were carried out in which solid cultivation of substrate was performed in an aerated packed-bed bioreactor for 7 days. Experimental results showed that the highest activity of β-xylosidase (6.13 U g-1) was obtained at an incubation, an initial moisture level and an aeration rate of 32.5°C, 60% and 1.5 L min-1, respectively. Statistical analysis revealed that the quadratic terms of incubation temperature and initial moisture content had highly significant effects on the production of β-xylosidase (p<0.01). Optimum conditions suggested by the statistical model for the production of β-xylosidase were 32.3°C incubation temperature, 59.7% initial moisture and 0.5 L min-1 aeration rate for attaining a predicted production level of 5.32 U g-1.
PDF Abstract XML References Citation
How to cite this article
P. Abdeshahian, N. Samat and W.M. Wan Yusoff, 2010. Production of β-Xylosidase by Aspergillus niger FTCC 5003 Using Palm Kernel Cake in a Packed-Bed Bioreactor. Journal of Applied Sciences, 10: 419-424.
DOI: 10.3923/jas.2010.419.424
URL: https://scialert.net/abstract/?doi=jas.2010.419.424
DOI: 10.3923/jas.2010.419.424
URL: https://scialert.net/abstract/?doi=jas.2010.419.424
INTRODUCTION
Xylan is the main constituent of hemicellulose and a major structural component of plant cell walls (Sá-Pereira et al., 2003). Xylans are composed of a linear backbone of β-1, 4-linked D-xylopyranosyl units, which may contain different side chains. β-xylosidase (EC 3.2.1.37) is a xylanolytic enzyme that hydrolyzes the non-reducing ends of xylooligosaccharides to release xylose and to catalyze the cleavage of xylobiose (Pedersen et al., 2007).
Several tropical agro-industrial residues have been used to produce value-added products particularly industrial enzymes (Pandey et al., 2001). Palm Kernel Cake (PKC) is a solid residue of palm oil industry in tropical area that is mostly used as a supplementary feed for farm animals (Ong et al., 2004).
The bioprocess of agro-industrial residues in Solid Substrate Fermentation (SSF) has been found to be a suitable method for the production of microbial enzymes such as cellulase, xylanase and β-xylosidase. The SSF is defined as the growth of microorganisms on insoluble materials in absence or near absence of free water (Pandey et al., 2001). Solid substrate fermentation process can be affected by various environmental variables such as temperature, moisture content of substrate and airflow. These variables are able to influence product formation process (Pandey et al., 1999).
A series of studies have been carried out on the production of β-xylosidase using hemicellulosic substances in SSF process. However, most of the investigations have aimed at producing of enzyme in shake flasks and a little information is available about the utilization of bioreactors for the production of β-xylosidase in SSF process (Poutanen and Puls, 1988; Pedersen et al., 2007; Rajoka, 2007; Mamma et al., 2008). A number of studies have been undertaken to utilize PKC in the production of industrial enzymes such as α-amylase, metalloprotease, tannase and β-mannanase (Abdeshahian et al., 2010; Ramachandran et al., 2004; Sabu et al., 2005; Sumantha et al., 2005). The aim of present study was to evaluate the production of β-xylosidase by Aspergillus niger FTCC 5003 using PKC in a laboratory packed-bed bioreactor under SSF process. In this study the combined effects of incubation temperature, moisture content and aeration rate on β-xylosidase production was determine using Response Surface Methodology (RSM) (Box et al., 1978).
MATERIALS AND METHODS
Design of experiment: A number of experiments were designed based on a 23 full factorial, Central Composite Design (CCD) to generate eighteen treatment combinations for three independent variables and each variable varied at three levels. This design included 6 star points and 4 replicates at the central point. The distance from the center of the design space to a star point was +1 or -1 unit (Haaland, 1989). The experimental variables studied were incubation temperature, initial moisture content and aeration rate. Each variable was coded at three levels of -1, 0, +1 representing low, middle and high level of the variables, respectively. The design was represented by a second-order polynomial regression model according to the following equation:
| (1) |
where, Y is the measured response (β-xylosidase, U g-1), Xi and Xj are the independent variables, ao represents the intercept and ai, aii and aij are the regression coefficients of the model.
For three variables, the model equation is shown below:
| (2) |
where a1, a2 and a3 are linear coefficients, a11, a22 and a33 are squared coefficients and a12, a13 and a23 are interaction coefficients.
The coded values and the actual levels of the variables are given in Table 1. The statistical analysis of the data was performed using Design-Expert software (version 6.0.6 Stat-Ease, Inc.). The design of experiments is given in Table 2.
Microorganism: The laboratory strain FTCC 5003 of Aspergillus niger was used in this study. The stock culture was propagated on a Potato Dextrose Agar (PDA) slant at 30°C for 7 days. For inoculum preparation 50 mL of the sterile Tween-80 (0.1%) was added to a PDA slant and aliquots (1.0 mL) of the mixture were used to inoculate substrate.
Media and growth conditions: Finely ground palm kernel cake was sieved and particles (2 mm size) were dried in an oven at 60°C for 48 h.
| Table 1: | Independent variables and levels used in central composite design |
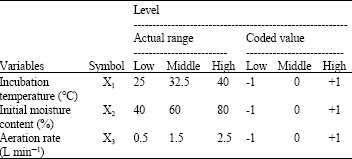 | |
| Table 2: | Central composite design and experimental results of β-xylosidase production by Aspergillus niger FTCC 5003 using palm kernel cake in SSF process for 7 days |
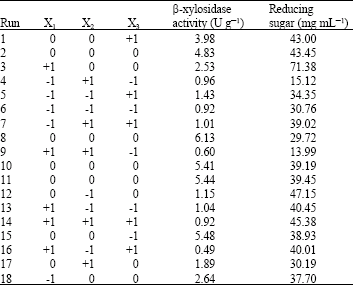 | |
| X1: Temperature (°C); X2: Moisture level (%); X3: Airflow rate (L min-1) | |
The total amount of 100 g of dried PKC was transfered to a laboratory packed-bed bioreactor (30 cm height and 16 cm inner diameter) and sterilized at 121°C for 30 min. PKC was moistened by Mandels’ medium (Sternberg, 1976) with levels according to experimental design. The composition of Mandels’ medium was as follows (g L-1): (NH4)2SO4, 1.4; KH2PO4, 2.0; CaCl2, 0.3; MgSO4.7H2O, 0.3; MnSO4.H2O, 0.0016; FeSO4.7H2O, 0.005; ZnSO4.7H2O, 0.0014; CoCl2, 0.002; protease peptone, 0.75; urea, 0.3; and Tween 80, 1.0. The initial pH of Mandels’ medium was adjusted to 5.0. Moistened PKC was inoculated with 1.0 mL of spore suspension containing 106 spores per gram of dry PKC. Culture was then incubated for 7 days under aeration and temperature determined by the experimental design.
Culture extraction: Enzyme extract was prepared by the addition of 1000 mL of 0.05 M sodium citrate buffer (pH 5.0) to fermented PKC and subsequent shaking at 170 rpm for 24 h under 4°C. Filter paper (Whatman No. 1) was used to separate culture residues from liquid supernatant at the temperature of 4°C. The filtered extract was used as the source of enzyme.
Enzyme assay: β-xylosidase activity was determined by the incubation of 0.2 mL of culture extract with 1.8 mL of 0.005 M solution of ρ-nitrophenyl-β-D-xylopyranoside prepared in 0.05 M sodium citrate buffer (pH 5.3). The reaction mixture was incubated at 50°C for 10 min. The reaction was stopped by the addition of 1.0 mL of 1.0 M sodium bicarbonate solution. The absorbance of liberated ρ-nitrophenol was determined at wavelength of 430 nm (Poutanen and Puls, 1988). One unit (U) of enzyme activity was defined as the amount of enzyme required to liberate 1.0 μmol of ρ-nitrophenol per min under the assay conditions. The activity of β-xylosidase produced was expressed in unit per gram of dry PKC (U g-1). Reducing sugar concentration was determined by dinitrosalicylic acid (DNS) method as described by Miller (1959).
RESULTS
Test results: Experimental results of β-xylosidase production based on CCD design are shown in Table 2. As observed, the highest β-xylosidase production (6.13 U g-1) was produced when Aspergillus niger (FTCC 5003) was grown at an incubation temperature of 32.5°C, an initial moisture of 60% and an aeration rate of 1.5 L min-1 (Run 8). As is evident, the peak level of reducing sugar (71.38 mg mL-1) was obtained in run 3. As shown in Table 2, the lowest production of β-xylosidase (0.49 U g-1) was measured at 40°C temperature, 40% moisture level and 2.5 L min-1 aeration rate (Run 16).
Regression model: By applying multiple regression analysis to the test results, a second-order polynomial Eq. 3 was obtained:
| (3) |
where, Y is β-xylosidase activity (U g-1) and X1, X2 and X3 are coded values of incubation temperature (oC), initial moisture content (%) and aeration rate (L min-1), respectively. The Analysis of Variance (ANOVA) was used to evaluate the significance of the fitted model (Table 3). As found, computed model F value of 15.49 with a probability value (Prob>F) 0.0004 revealed that the selected quadratic model was significantly fitted to the experimental data. The lack of fit related to F value (4.92) suggested that the lack of fit was not significant and hence model was fit to the data. The values of Prob>F less than 0.05 indicate that the model terms are significant. As observed, the quadratic terms of temperature (X12) and moisture content (X22) were significant at 99% probability level (p<0.01). The multiple coefficient of determination (R2) with a reasonable value of 0.9457 implied that 5.43% of sample variation was not attributed to the variables tested.
Response surface graph: The three dimensional response surface plots described by the regression model were drawn to illustrate the effects of the independent variables and the interactive effects of the independent variables on the response formed (Fig. 1-3). Figure 1 shows the effects of incubation temperature and initial moisture content of PKC on the production of β-xylosidase. As can be seen, the yield of β-xylosidase increased higher than 0.54 U g-1 when temperature began to increase from 25°C to optimum range (31-32.5°C) with the low level of moisture content. Following a rise in moisture content the level of β-xylosidase activity significantly increased to 4.92 U g-1. However, a decrease in response was observed when temperature increased more than optimum range. Similarly, β-xylosidase yield declined when the rise in initial moisture content was greater than optimum range (59-60%). Figure 2 shows the simultaneous influences of moisture level and aeration rate on β-xylosidase production.
| Table 3: | Analysis of variance for the quadratic model of β-xylosidase production by Aspergillus niger FTCC 5003 grown on PKC in SSF for 7 days |
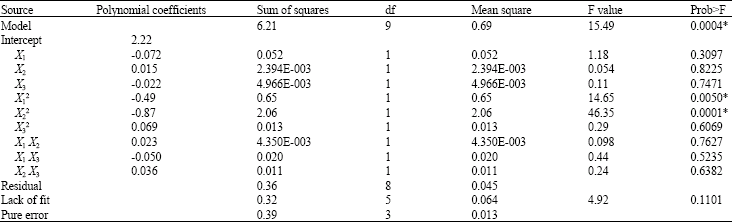 | |
| *Statistically significant at 99% probability level. X1: Temperature (°C); X2: Moisture level (%); X3: Airflow rate (L min-1). X12, X22 and X32: The quadratic terms; X1X2, X1 X3 and X2 X3: The interaction terms. R2 = 0.9457 | |
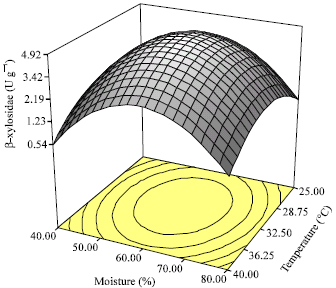 | |
| Fig. 1: | Three-dimensional response surface graph of the combined effects of incubation temperature and moisture content on β-xylosidase production by Aspergillus niger FTCC 5003 using palm kernel cake in SSF process for 7 days |
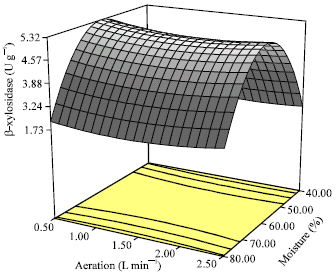 | |
| Fig. 2: | Three-dimensional response surface graph of the combined effects of moisture content and aeration rate on β-xylosidase production by Aspergillus niger FTCC 5003 using palm kernel cake in SSF process for 7 days |
Evidently, an increase in moisture content from 40% to optimum range concomitantly caused the considerable increase of β-xylosidase to maximum 5.32 U g-1, while an aeration rise higher than 0.5 L min-1 showed a slight decrease in β-xylosidase activity. Similar trend between temperature and aeration rate is shown in Fig. 3. As shown, no significant effect of aeration rate on β-xylosidase activity was observed at optimum temperature conditions.
 | |
| Fig. 3: | Three-dimensional response surface graph of the combined effects of incubation temperature and aeration rate on β-xylosidase production by Aspergillus niger FTCC 5003 using palm kernel cake in SSF process for 7 days |
Optimum conditions: The optimal levels of the variables for the production of β-xylosidase were determined by moving along the variable axis versus response obtained. The critical points presenting a maximum β-xylosidase activity had following values of 32.3°C, 59.7% and 0.5 L min-1 for incubation temperature, initial moisture content and aeration rate, respectively. The predicted β-xylosidase activity for these conditions was 5.32 U g-1.
DISCUSSION
Incubation temperature: The current study demonstrated that the increase of incubation temperature from 25°C to optimum level (32.3°C) raised the production of β-xylosidase up to the highest activities (Fig. 1, 3). Similar to our study, Panagiotou et al. (2003) showed that the peak level of β-xylosidase produced by Fusarium oxysporum on corn stover was measured when the growth temperature was set at 30°C. They observed that the activity of β-xylosidase produced at the temperature of 30°C was similar to that at 33°C, whereas it was higher than that at the temperature of 27°C. By contrast, Kalogeris et al. (2003) showed the peak level of β-xylosidase produced by Thermoascus aurantiacus on wheat straw was obtained at the temperature of 49°C. The variation in incubation temperature can be attributed to this fact that the elevated temperature may affect the membrane of vesicles in the filamentous fungi. Therefore, temperature changes in membrane structure leads to initiation of metabolic change and product formation (Tao et al., 1997). As can be seen from Fig. 1 and 3, the activity of β-xylosidase decreased when temperature increased greater than optimum level of 32.3°C. High temperature causes the reduction of microbial growth and metabolic activity (Venkateswarlu et al., 2000; Pandey et al., 2001). The production of β-xylosidase, therefore, decreased from 32.3 to 40°C.
Initial moisture content: As shown in Fig. 1 and 2, the production of β-xylosidase was remarkably dependent on moisture content so that the increase of moisture level from 40% to optimum level (59.7%) resulted in a quadratic rise in the production of β-xylosidase (Fig. 1, 2). The low level of moisture content causes the reduction of the growth of microorganism which consequently results in an decrease in enzyme production (Prior et al., 1992; Venkateswarlu et al., 2000). The current study showed that the increase of moisture level from 40% to optimum level (59.7%) improved the production of β-xylosidase. The increase of moisture favors the growth of microorganism. As a consequence, the production of enzyme enhances in high moisture (Gao et al., 2008). However, a drop in the activity of β-xylosidase was observed with too much moisture content (70-80%). This result could be attributed to the reductions of heat and mass transmission through the culture and the decrease of air exchange. Contrary to the results obtained in the present study, Mamma et al. (2008) showed that the growth of Aspergillus niger on Water Unextractable Orange Peel (WUOP) with initial moisture 90% led to the highest yield of β-xylosidase in culture. Evidently, high moisture level (79%) was found to be optimum level for maximal production of β-xylosidase when Fusarium oxysporum was grown on the mixture of brewer’s spent grain and corn cobs under SSF process (Xiros et al., 2008). The variations in optimum moisture content could be related to water-binding characteristics of substrate, temperature and selected microorganism (Prior et al., 1992).
Aeration rate: Figure 2 and 3 show that a rise in airflow rate (more than 0.5 L min-1) had no favorable effect on β-xylosidase production which was possibly due to deleterious effect of high aeration on the morphology of filamentous fungi and the decrease of moisture content (Lu et al., 1997; Mitchell et al., 1999). However, increasing aeration rate from 1 to 15 L min-1 led to an increase in the production of β-xylosidase by Thermoascus aurantiacus on wheat straw (Kalogeris et al., 2003). These differences could be related to the selection of microorganism, the particular amount of oxygen for product synthesis and the level of heat evolution to be removed (Lonsane et al., 1985).
CONCLUSION
The findings of this study reveal that PKC is able to induce β-xylosidase in SSF process using Aspergillus niger as a potential enzyme producer. According statistical model used, incubation temperature and initial moisture content had significant effect on β-xylosidase production in the packed-bed bioreactor.
ACKNOWLEDGMENTS
The authors would like to thank Dr. Barry McClear (Megazyme International Ireland Ltd.) for providing chemicals. Authors also thank Malaysian Agriculture Research and Development Institute (MARDI) for providing equipments and facilities during experimental study.
REFERENCES
- Abdeshahian, P., N. Samat, A.A. Hamid and W.M.W. Yusoff, 2010. Utilization of palm kernel cake for production of β-mannanase by Aspergillus niger FTCC 5003 in solid substrate fermentation using an aerated column bioreactor. J. Ind. Microbiol. Biotechnol., 37: 103-109.
CrossRef - Gao, J., H. Weng, D. Zhu, M. Yuan, F. Guan and Y. Xi, 2008. Production and characterization of cellulolytic enzymes from the thermoacidophilic fungal Aspergillus terreus M11 under solid-state cultivation of corn stover. Bioresour. Technol., 99: 7623-7629.
CrossRefDirect Link - Kalogeris, E., F. Iniotaki, E. Topakas, P. Christakopoulos, D. Kekos and B.J. Macris, 2003. Performance of an intermittent agitation rotating drum type bioreactor for solid-state fermentation of wheat straw. Bioresour. Technol., 86: 207-213.
CrossRef - Lonsane, B.K., N.P. Ghildyal, S. Budiatman and S.V. Ramakrishna, 1985. Engineering aspects of solid state fermentation. Enzyme Microbiol. Technol., 7: 258-265.
CrossRef - Lu, M., J.D. Brooks and I.S. Maddox, 1997. Citric acid production by solid-state fermentation in a packed-bed reactor using Aspergillus niger. Enzym. Microb. Technol., 21: 392-397.
CrossRef - Mamma, D., E. Kourtoglou and P. Christakopoulos, 2008. Fungal multienzyme production on industrial by-products of the citrus-processing industry. Bioresour. Technol., 99: 2373-2383.
CrossRefPubMedDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Mitchell, D.A., A. Pandey, S. Penjit and N. Krieger, 1999. Scale-up strategies for packed-bed bioreactors for solid-state fermentation. Process. Biochem., 35: 167-178.
Direct Link - Ong, L.G.A., S. Abd-Aziz, S. Noraini, M.I.A. Karim and M.A. Hassan, 2004. Enzyme production and profile by Aspergillus niger during solid substrate fermentation using palm kernel cake as substrate. Applied Biochem. Biotechnol., 118: 73-79.
Direct Link - Panagiotou, G., D. Kekos, B.J. Macris and P. Christakopoulos, 2003. Production of cellulolytic and xylanolytic enzymes by Fusarium oxysporum grown on corn stover in solid state fermentation. Ind. Crops. Prod., 18: 37-45.
CrossRefDirect Link - Pedersen, M., H.K. Lauritzen, J.C. Frisvad and A.S. Meyer, 2007. Identification of thermostable b-xylosidase activities produced by Aspergillus brasiliensis and Aspergillus niger. Biotechnol. Lett., 29: 743-748.
Direct Link - Poutanen, K. and J. Puls, 1988. Charactristic of Trichoderma reesei: β-xylosidase and its use in the hydrolysis of solublized xylans. Applied Microbiol. Biotechnol., 28: 425-432.
CrossRef - Rajoka, M.I., 2007. Kinetic parameters and thermodynamic values of β-xylosidase production by Kluyveromyces marxianus. Bioresour. Technol., 98: 2212-2219.
CrossRef - Ramachandran, S., A.K. Patel., K.M. Nampoothiri, S. Chandran, G. Szakacs, C.R. Soccol and A. Pandey, 2004. α-amylase from a fungal culture grown on oil cakes and its properties. Braz. Arch. Biol. Technol., 47: 309-317.
Direct Link - Sa-Pereira, P., H. Paveia, M. Costa-Ferreira and M.R. Aires-Barros, 2003. A new look at xylanase: An overview of purification strategies. Mol. Biotechnol., 24: 257-281.
Direct Link - Sabu, A., A. Pandey, M.J. Daud and G. Szakacs, 2005. Tamarind seed powder and palm kernel cake: Two novel agro residues for the production of tannase under solid state fermentation by Aspergillus niger ATCC 16620. Bioresour. Technol., 96: 1223-1228.
Direct Link - Sternberg, D., 1976. Production of cellulase by Trichoderma. Biotechnol. Bioeng. Symp., 6: 35-53.
Direct Link - Tao, S., L. Beihui, L. Deming and L. Zuohu, 1997. Effect of elevated temperature on Trichoderma viride SL-1 in solid state fermentations. Biotechnol. Lett., 19: 171-174.
Direct Link - Venkateswarlu, G., P.S.M. Krishna, A. Pandey and L.V. Rao, 2000. Evaluation of Amycolatopsis mediterranei VA18 for production of rifamycin-B. Process. Biochem., 36: 305-309.
Direct Link - Xirosa, C., E. Topakasa, P. Katapodisa and P. Christakopoulos, 2008. Evaluation of Fusarium oxysporum as an enzyme factory for the hydrolysis of brewer's spent grain with improved biodegradability for ethanol production. Industrial Crops Prod., 28: 213-224.
CrossRefDirect Link