
S.M.N. Amin
Institute of Marine Sciences and Fisheries, University of Chittagong, Chittagong-4331, Bangladesh
A. Arshad
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
J.S. Bujang
Department of Animal Science and Fishery, Faculty of Agriculture and Food Sciences, Universiti Putra Malaysia, Bintulu Campus, 89007 Bintulu Sarawak, Malaysia
S.S. Siraj
Department of Aquaculture, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
Journal of Applied Sciences
Year: 2009 | Volume: 9 | Issue: 5 | Page No.: 801-814







ABSTRACT
Age structure, growth, mortality and yield-per-recruit of Acetes indicus were examined in the coastal waters of Malacca, Peninsular Malaysia between February 2005 and January 2006. Monthly length frequency data were analyzed using FiSAT software for estimating population parameters, including asymptotic length (L∝), growth co-efficient (K) and exploitation rate (E) to assess the status of the stock. The L∝ and K for males were estimated at 29.40 mm and 1.70 year-1 and for the females that were 42 mm and 1.20 years-1, respectively. The growth performance index (φ’) was calculated as 3.16 and 3.33 for males and females. The growth pattern of males and females showed positive allometric nature of growth (b>3, p<0.05). The maximum life span (tmax) of males and females was 1.76 and 2.50 years, respectively. Total mortality (Z) by length converted catch curve was estimated at 4.15 year-1 for males and 3.50 year-1 for females. The rate of natural mortality (M) for males and females was calculated as 2.65 and 1.91 year-1 and the fishing mortality (F) was 1.50 and 1.59 year-1 for males and females, respectively. The recruitment pattern of A. indicus was continuous throughout the year with two major peaks. The exploitation rate (E) of males was 0.36 and that of females was 0.45. The maximum allowable limit of exploitation (Emax) of males and females was 0.71 and 0.57 for the highest yield. The exploitation rates were less than the predicted Emax values of males and females. Thus, the stock of A. indicus was found to be under exploited in the investigated area.
PDF Abstract XML References Citation
How to cite this article
S.M.N. Amin, A. Arshad, J.S. Bujang and S.S. Siraj, 2009. Age Structure, Growth, Mortality and Yield-Per-Recruit of Sergestid Shrimp, Acetes indicus (Decapoda: Sergestidae) From the Coastal Waters of Malacca, Peninsular Malaysia. Journal of Applied Sciences, 9: 801-814.
DOI: 10.3923/jas.2009.801.814
URL: https://scialert.net/abstract/?doi=jas.2009.801.814
DOI: 10.3923/jas.2009.801.814
URL: https://scialert.net/abstract/?doi=jas.2009.801.814
INTRODUCTION
The shrimp of the genus Acetes, is mainly used in subsistence fisheries and therefore, commercially important in the Peninsular Malaysia (Amin et al., 2008). Knowledge on the shrimp’s population characteristics are important if proper management of those resources is to be effective. Age in crustacean is difficult to estimate because exoskeletons are lost during moulting process (Xiao and Greenwood, 1993). Thus, analysis of length-frequency data has been used to identify year classes (Amin and Zafar, 2003, 2004; Amin et al., 2008; Paraconstantinou and Kapiris, 2001). Length-weight relationships are useful for a wide number of studies, such as estimating population structure (Smith et al., 2008), growth rates and other aspects of fish/shrimps population dynamics (Tsoumani et al., 2006).
The geographical distribution of Acetes has been described by Xiao and Greenwood (1993). This species A. indicus occurs in the central part of the Indo-West Pacific. The distribution ranges from the South China Sea (Dong Hoa) through the Gulf of Siam (Sanudpragarn), Java (Surabaya), Straits of Malacca (Singapore and Malacca) and the Bay of Bengal to the Arabian Sea. The shrimp plays a substantial role in the food webs of coastal waters, acting as predators, feeding on a variety of foods ranging from diatoms, copepods and larvae of decapods to detritus and in turn as prey for many fishes and other predators (Xiao and Greenwood, 1993). It appears in very large swarms in the shallow inshore coastal waters, which is brackish with a salinity of 30 ppt or less, during certain seasons of the year. In West coast of Malay Peninsula, A. indicus, along with A. japonicus, is commercially exploited from March to November, using the estuarine push net (Arshad et al., 2007).
Spectacular school or swarms of Acetes, particularly in coastal waters of Asia, are the bases of important commercial fishes for consumption by humans and domestic animals (Omori, 1975, 1978). The commercial importance also derives from the use and potential of Acetes as a food organism for aquaculture industry (Kungvankij et al., 1986). These combined features make Acetes excellent candidates for population dynamics studies. In spite of greater abundance and importance of Acetes in the fishery of Asian countries, very little information is available on the population dynamics so far except the studies carried out by Zafar et al. (1997, 1998), Zafar and Amin (2002) and Oh and Jeong (2003).
Knowledge of various population parameters like as asymptotic length (L∝), growth coefficient (K), motilities (natural and fishing) rate and exploitation level (E) are necessary for planning and management of Acetes resources. There are many tools for assessing exploitation level and status of stock. Of these, FiSAT (FAO-ICLARM Stock Assessment Tools) has been most frequently used for estimating population parameters of shrimps (Jayawardane et al., 2002, 2003; Paraconstantinou and Kapiris, 2001; Etim and Sankare, 1998; Enin et al., 1996) primarily because it requires only length-frequency data. The objective of the present study was to estimate the population parameters and exploitation level of A. indicus in order to assess the stock status of the species around the coast waters of Malacca, Peninsular Malaysia and to provide data that could be useful for management.
MATERIALS AND METHODS
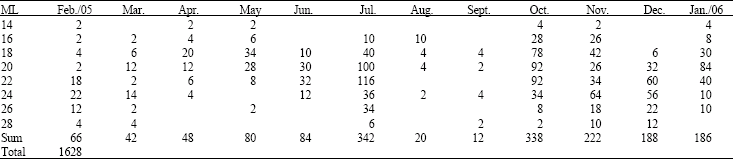
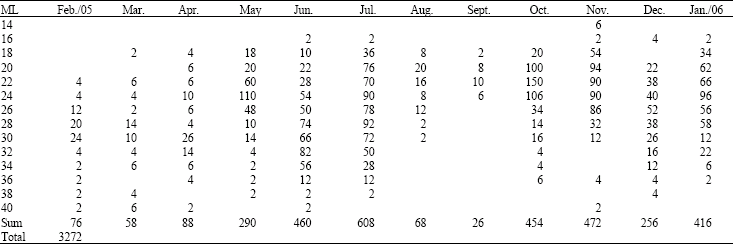
Collection of data: Monthly fresh samples of A. indicus were collected between February 2005 and January 2006 from commercial push net catches landed at Klebang Besar (N 02° 13.009’ and E 102° 11.921’), Malacca, Peninsular Malaysia (Fig. 1). Acetes shrimps were caught by the push net (triangular shape) known locally as Sungkor, as described by Omori (1975) in the coastal waters of Klebang Besar, Malacca, Peninsular Malaysia. Dimensions of the net were 5-6 m in length, 4.0-4.5 m in wide and 3.0 -3.5 m in height. The mean mesh sizes were 3.2 ± 0.27 cm at the anterior section, 0.75 ± 0.0 cm at the middle and 0.5 ± 0.08 at cod end (stretched). After collection, samples were fixed in 10% formalin solution in the field and analyzed after 2-3 days of preservation. In the laboratory, specimens were identified using a Nikon dissecting microscope (Nikon-122764, Japan). Sexes were determined by the presence or absence of petasma on the first pleopods and clasping spine on the lower antennular flagellum (Omori, 1975). The studies of Omori (1975) were followed during the identification of different species of Acetes. Total Length (TL) of 4900 individuals (males 1628 and females 3272) was measured from the tip of the rostrum to the tip of the telson to the nearest 0.1 mm and total weight was taken by an electronic balance of 0.001 g accuracy. The length-frequency distributions for males and females A. indicus samples are shown in Table 1 and 2.
Data analysis: To estimate the age structure, the length-frequency data of A. indicus were analyzed by using the MINITAB Version 14 and SPSS Version 11.5. The differences in the size-frequency distributions of population between sexes were determined by the Kolmogorov-Smirnov two-sample test (Sokal and Rohlf, 1995). Student’s t-test was used for comparison of the mean total length of males and females (Zar, 1996). Size-frequency distributions of A. indicus were plotted for each month from February 2005 to January 2006. Bhattacharya’s method, implemented from the package FiSAT (Gayanilo et al., 1996), was used to identify the modes in the polymodal length-frequency distributions of A. indicus. All the identified size/age groups were derived from at least three consecutive points and selection of the best results was based on the following criteria: (a) the values of Separation Index (SI) for the different age groups; (b) No. of the identified age groups and (c) Standard Deviation (SD) (Gayanilo et al., 1989).
To establish the length-weight relationship, W = a Lb was applied (Quinn II and Deriso, 1989), where, W is the weight (mg), L is the total length (mm), a is the intercept (condition factor) and b is the slope (relative growth rate). The parameters a and b were estimated by least squares linear regression on log-log transformed data: Log10 W = Log10 a+b Log10 L.
| Table 1: | Length-frequency data of male A. indicus from the coastal waters of Malacca, Peninsular Malaysia |
 | |
| Table 2: | Length-frequency data of female A. indicus from the coastal waters of Malacca, Peninsular Malaysia |
 | |
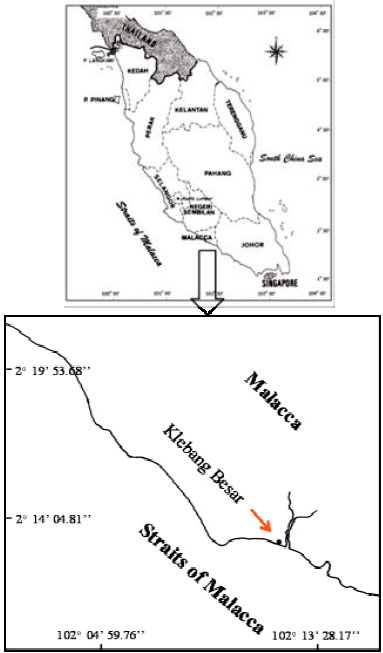 | |
| Fig. 1: | Sampling station (dot) and location of Klebang Besar, Malacca, Peninsular Malaysia |
The coefficient of determination (r2) was used as an indicator of the quality of the linear regression (Scherrer, 1984). Additionally, 95% confidence limits of the parameter b and the statistical significance level of r2 were estimated.
Monthly length-frequency distributions of male and female A. indicus were analyzed using the FiSAT computer programme (Gayanilo et al., 1996). An estimate of maximum length (Lmax) was obtained using the data described earlier and the extreme value theory (Formacion et al., 1991) as implemented in the FiSAT software. The parameters of the von Bertalanffy Growth Function (VBGF), asymptotic length (L∝) and growth co-efficient (K) were estimated by means of ELEFAN-I (Pauly and David, 1981). Estimated of L∝ and K was used to calculate the growth performance index (φ’) (Pauly and Munro, 1984) using the equation:
φ’ = 2 log10L∝ + log10K |
Potential longevity (tmax) of the species was calculated from Pauly (1984) formula: tmax = 3/K. The inverse von Bertalanffy growth equation (Sparre and Venema, 1992) was used to determine male and female of A. indicus lengths at various ages. The VBGF is defined by the equation:
Where:
| Lt | = | Mean length at age t |
| L∝ | = | Asymptotic length |
| K | = | Growth co-efficient |
| t | = | Age of the A. indicus |
| t0 | = | The hypothetical age at which the length is zero (Dulcic and Kraljevic, 1995) |
Once the growth parameters of VBGF were obtained, a linearized length-converted catch curve was constructed using the following formula to estimate total mortality (Z):
In (Nt/Δt) = a+bt |
where, N is the No. of individuals of relative age (t) and Δt is the time needed for the shrimp to grow through a length class. The slope (b) of the curve with its sign changed gives Z.
Natural mortality (M) was estimated using the empirical relationship of Pauly (1980):
Log10M = - 0.0066 – 0.279Log10L∝ + 0.6543.Log10K + 0.4634 Log10T |
where, M is the natural mortality, L∝ the asymptotic length, K the growth co-efficient of the VBGF and T the mean annual habitat water temperature °C (here it was 30.58 °C).
Once Z and M were obtained, fishing mortality (F) was estimated using the relationship:
F = Z – M |
where, Z is the total mortality and M, natural mortality. The exploitation level (E) was obtained from E = F/Z (Gulland, 1971).
Ascending left arm of the length-converted catch curve was used to estimate the probability of capture. By plotting the cumulative probability of capture against mid-length of class interval, we obtain a resultant curve from which the length at first capture Lc was taken as corresponding to the cumulative probability at 50%.
The recruitment pattern was obtained by projecting the length-frequency data backwards on the time axis using growth parameters (Moreau and Cuende, 1991). Normal distribution of the recruitment pattern was determined by NORMSEP (Pauly and Caddy, 1985) in FiSAT.
Relative yield-per-recruit (Y/R) and relative biomass-per-recruit (B/R) were estimated according to Beverton and Holt (1966) length-based method as modified by Pauly and Soriano (1986) using the knife-edge selection. From the analysis, Emax (maximum exploitation rate giving maximum relative yield-per-recruit) and E0.5 (the value of exploitation rate at 50% of the unexploited relative biomass-per-recruit) were estimated.
RESULTS
Size frequency distribution: Between February 2005 and January 2006, total number of individuals collected for this study were 1628 (33.22%) for males and 3272 (66.78%) for females, respectively. In males, the minimum and maximum total lengths were 13 and 29 mm and in females, they were 13 and 41 mm, respectively. The mean total lengths were 21.37 ± 2.86 and 25.44 ± 4.66 mm for males and females, respectively (Fig. 2). According to the annual size-frequency distributions, there were significant difference between males and females (Kolmogorov-Smirnov test: dmax = 0.396, p<0.001). The mean total length of females was 4.08 mm taller than that of the males (Fig. 3) and it was significantly different (t-test, p<0.001). Monthly size frequency distribution over 12 months suggested that the population consisted of a maximum of two age groups (Fig. 4) with mean values being 21.01 and 30.98 mm of total length.
| Table 3: | Estimated length-weight parameters of A. indicus from the coastal water of Malacca, Peninsular Malaysia |
 | |
| N: Sample size; TL range: Minimum and maximum total length (mm); TW range: Minimum and maximum total weight (mg); a and b: Parameters of the length-weight relationship; CI: Confidence Interval; r2: Coefficient of determination | |
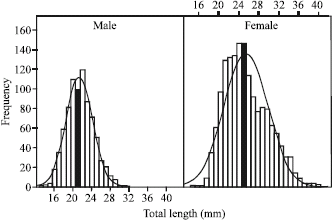 | |
| Fig. 2: | Annual length-frequency distribution of males and females A. indicus collected from the coastal waters of Malacca, Peninsular Malaysia |
 | |
| Fig. 3: | Box plots of male and female Acetes indicus from the coastal waters of Malacca |
Length-weight relationships: Length and weight of individuals for determining the length-weight relationships ranged from 13-29 mm and 9.50-174.10 mg for males, 13 to 41 mm and 13.30 to 430 mg for females, respectively (Table 3). The regression between TL (total length) and TW (total weight) for the males and females showed positive relationship (Fig. 5). The parameters of total length-body weight relationship for different groups (males, females and combined sexes) are shown in Table 3 and the length-weight relationship equations were established as:
| Table 4: | Estimated population parameters of A. indicus from the coastal waters of Malacca, Peninsular Malaysia |
 | |
Log TW = 3.307 Log TL – 2.7136, r2 = 0.85 for male A. indicus
Log TW = 3.472 Log TL – 2.9437, r2 = 0.92 for female A. indicus
Log TW = 3.411 Log TL – 2.8565, r2 = 0.92 for combined sexes of A. indicus
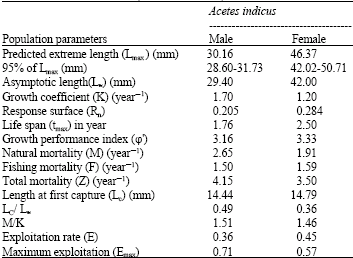
Growth parameters: The observed length and predicted extreme length (Lmax) of males were 28.00 and 30.16 mm, respectively with 95% confidence interval for extreme length was 28.60-31.73 mm. For females the observed length and predicted extreme length (Lmax) were 40 and 46.37 mm, respectively with 95% confidence interval for extreme length was 42.02 to 50.71 mm (Table 4). The von Bertalanffy Growth Function (VBGF) parameters L ∝ and K in males were obtained for the best fit with L ∝ = 29.40 mm and K = 1.70 year-1 (Fig. 6a). For females, these parameters were estimated L ∝ = 42 mm and K = 1.20 year-1 (Fig. 6b). The response surface, Rn values were calculated by ELEFAN-I as 0.205 for males and 0.284 for females. The growth performance index (φ’) of male was 3.16 and that of females was 3.33.
Age and growth: Based on the growth coefficients, the maximum life span (tmax = 3/K) of males was 1.76 years and that of females was 2.50 years.
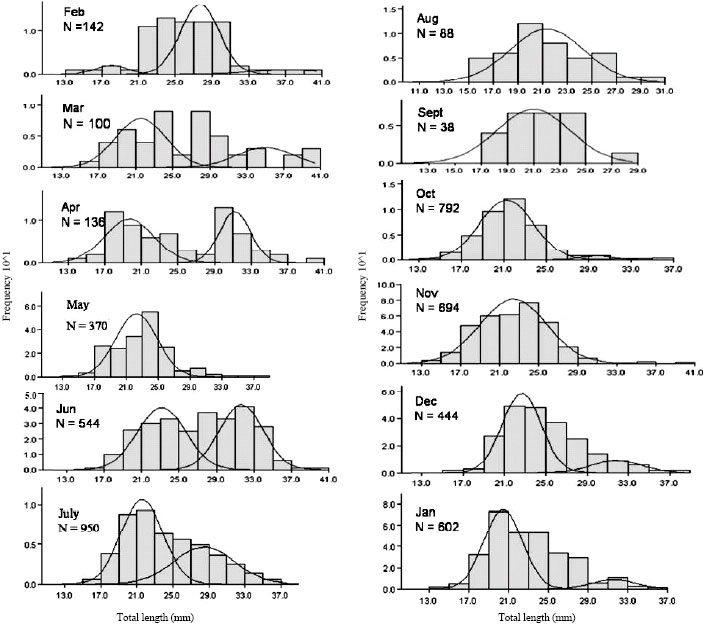 | |
| Fig. 4: | Monthly length frequency distributions of males and females A. indicus caught in the coastal waters of Malacca, Peninsular Malaysia |
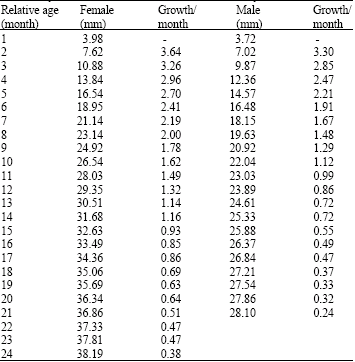
The application of modal progression analysis by Bhattacharya’s method determined modal lengths for combined sexes of A. indicus between 17 mm in February and 35 mm in March, with satisfactory separation index for combined sexes of A. indicus (Table 5). Maximum two well-discriminated modal groups were identified reflecting different annual cohorts (Table 5). Table 6 shows the total length-age relationship estimated from Bertalanffy’s model using L ∝ and K parameters of males and females and assuming t0 = 0 (Pauly and David, 1981). Therefore, the sizes attained by males and females A. indicus were 23.89 and 28.99 mm at the end of one year age, respectively (Table 6).
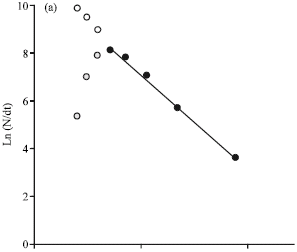
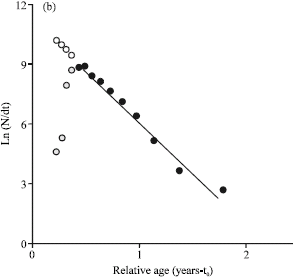
Mortality and exploitation: Total mortality (Z) was estimated using the length converted catch curve (Fig. 7).
The dark points of the curve were used by fitting a least square regression line to them. Good fit to the descending right hand limits of the catch curve was considered with mentioned regression line values:
| Male | : Y = 11.225 + (– 5.312) X (r = – 0.99) |
| Female | : Y = 10.398 + (– 4.778) X (r = – 0.99) |
The estimated total mortality, Z values were 4.15 and 3.50 year-1 for males (Fig. 7a) and females (Fig. 7b), respectively. The natural mortality rate, M values of males was 2.65 year-1 and that of females was 1.99 year-1. The fishing mortality rate (F) of males and females was 1.50 and 1.59 year-1, respectively (Table 4). The natural mortality of males (2.65 year-1) was higher than the value of females (1.99 year-1) indicating the unbalance position in the stock.
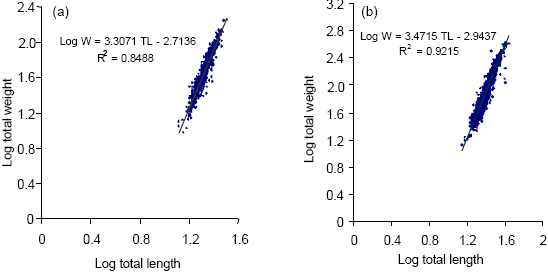 | |
| Fig. 5: | Length weight relationship of male (a) and female (b) A. indicus from the coastal water of Malacca, Peninsular Malaysia |
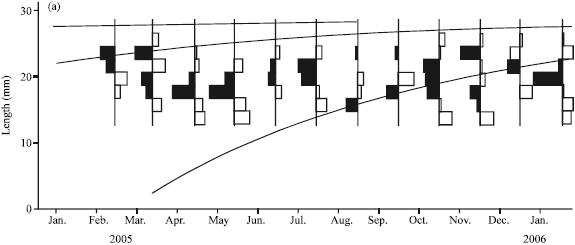 | |
| Fig. 6: | Von Bertalanffy growth curves of males (a) and females (b) A. indicus superimposed on the restructured length-frequency histograms. The black and white bars are positive and negative deviation from the weighted moving average of three length classes and they represent psedo-cohorts |
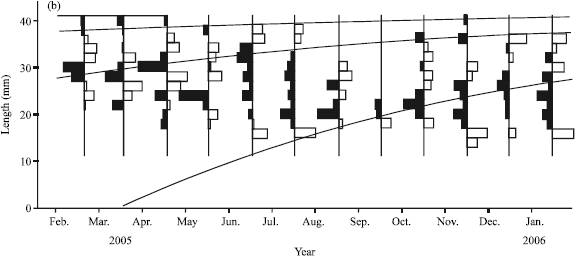
From these figures, exploitation rate, E values were obtained 0.36 and 0.45 for males and females in the coastal waters of Malacca, which seemed to be below to the optimum level of exploitation (E = 0.50).
| Table 5: | Identified age groups from the length-frequency analysis of male and female A. indicus during the monthly sampling, using Bhattacharya’s method |
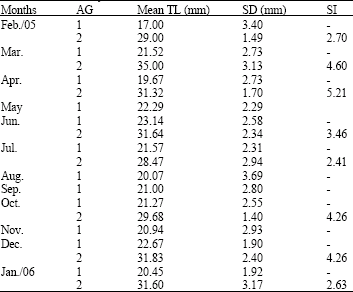 | |
| AG: Age Group, TL: Total Length, SD: Standard Deviation, SI: Separation Index | |
| Table 6: | Length-at-age key for A. indicus from coastal waters of Malacca, estimated using the VBGF parameters (L ∝ = 29.40 mm, K = 1.70 year-1 for males and L ∝ = 42 mm, K = 1.20 year-1) |
 | |
Length at first capture: The length at first capture, Lc (the length at which 50% of the shrimps becomes vulnerable to the gear) of A. indicus in the coastal waters of Malacca are shown in Fig. 8. The mean Lc value of males and females A. indicus was 14.44 and 14.79 mm, respectively during the study period. The L25% value of males and females was 12.35 and 12.04 mm. The mean length at which 75% of the shrimps were retained in the gear was estimated as L75% = 16.31 mm for males and L75% = 17.92 mm for females in the investigated area.
 | |
| Fig. 7: | Length converted catch curve of males (a) and females (b) A. indicus; the darkened full dots represent the points used in calculating through least square linear regression and the open dots represent the point either not fully recruited or nearing to L ∝ |
Recruitment pattern: The recruitment pattern of males was continuous throughout the year, with two major peaks (Fig. 9a). The first spell of males was from February to May with peak in March (11.59%) and the second spell was from July to October with peak in September (13.91%). In case of females, the recruitment pattern was also two spells during the 1-year study (Fig. 9b). The first spell of females was from January to March with peak in January (5.12%). The second spell was August to October with peak in September (21.32%).
Relative yield per recruit (Y/R) and biomass per recruit (B/R): The computed maximum allowable limit of exploitation (Emax) that give the maximum relative yield-per-recruit at maximum sustainable yield (MSY) level was 0.71 for males (Fig. 10a) and 0.57 for females (Fig. 10b).
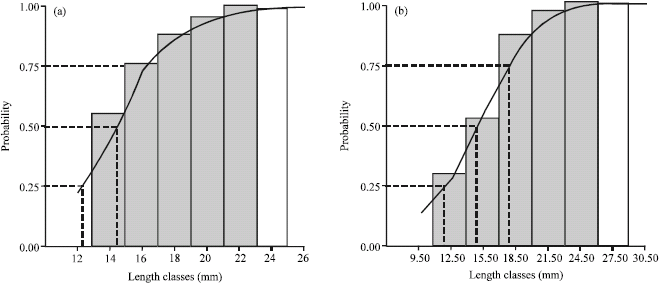 | |
| Fig. 8: | Probability of capture of each length class of males (a) and females (b) A. indicus (L25% = 12.35 mm, L50% or Lc = 14.44 mm and L75% = 16.31 mm for males and L25% = 12.04 mm, L50% or Lc = 14.79 mm and L75% = 17.92 mm for females) |
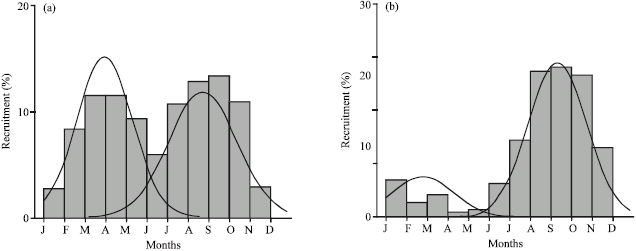 | |
| Fig. 9: | Recruitment pattern of males (a) and females (b) A. indius in the coastal waters of Malacca, Peninsular Malaysia |
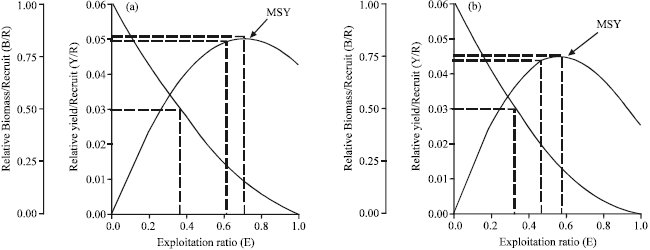 | |
| Fig. 10: | Yield-per-recruit and biomass per recruit models, showing levels of yield index of males (a) and females (b) A. indicus: MSY = Maximum Sustainable Yield |
The exploitation level (E-0.5) which corresponds to 50% of the relative biomass-per-recruit of the unexploited stock was 0.36 for males that of females was 0.32, respectively.
DISCUSSION
The size structure of the population in the investigated area consists of a relatively higher percentage of the females than the males. The females attained a greater size indicating a size dimorphism. The growth coefficient b generally lies between 2.5 and 3.5 and the relation is said to be isometric when it is equal to 3 reported for most aquatic organisms (Ecoutin et al., 2005). Table 7 shows earlier published values of the coefficient a and b for the genus Acetes. The values of b show considerable variation, ranging from 2.155 (Lei, 1988) to 3.472 (present study). Estimated b (3.307) of males lies between the values mentioned by Ecoutin et al. (2005) and is significantly higher than isometric value (3) at 5% level. In females, estimated b (3.472) is also significantly higher from then the isometric value (3) at 5% level. This indicates the positive allometric nature of growth for A. indicus in the coastal waters of Malacca.
Estimated L ∝ and K are 29.40 mm and 1.70 year-1 for males. In females they are 42 mm and 1.20 year-1. Comparisons with population parameters from other studies (Table 8) show that differences exist for different species of the genus Acetes from different areas in the world. The highest value of L ∝ (42 mm in females) is observed in the present study; the lowest value (31 mm in combined sexes) was in Bangladesh waters (Zafar et al., 1997). The K values are more or less similar to the present value except the reported by Oh and Jeong (2003) stated the lowest values (K = 0.84 year-1 in males and K = 0.69 year-1 in females for A. chinensis. The index of phi prime by Munro and Pauly (1983) is suitable for comparing and computing the overall growth performance of different species of shrimp stocks. The phi prime for this species with the present estimates of L ∝ and K were found to be excellent and varied between 3.16 and 3.33 (Table 8). Though phi prime is supported to be more or less constant for a family or for similar taxa, the range here (Table 8) is low except the report by Oh and Jeong (2003) stated the lowest values (φ’ = 1.97 in males and φ’ = 2.10 in females for A. chinensis).
Longevity of Acetes, as in other crustaceans, is difficult to estimate accurately because of the obvious difficulties in ageing these animals and hence estimates are unavailable for both natural and laboratory populations (Xiao and Greenwood, 1993). The maximum age, tmax = 3/K (Pauly, 1984) is frequently using for estimating longevity of shrimp (Botter-Carvalho et al., 2007; Niamaimandi et al., 2007; Cha et al., 2004). The estimated maximum age for males and females are 1.76 and 2.50 years in the present study. The mean life span of penaeids is often regarded as <2 year (Dall et al., 1990). Estimated longevity of Acetes is less than that of penaeids is well justified because of its smaller size. Yoshida (1949) suggested that Acetes chinensis has a life-span of about one year in western Korea. Except this no reports are available for comparison of longevity of Acetes so far researcher aware. Maximum of two age groups of combined sexes of A. indicus with mean values being 21.01 and 30.98 mm of total length were estimated by Bhattacharya’s method. Similar studies were reported on red shrimp Aristeus antennatus by Paraconstantinou and Kapris (2001) and on C. madrasensis by Amin et al. (2008), by the Bhattacharya’s method, which also followed in the present study.
| Table 7: | Parameters of length-weight relationship (a and b) for the genus Acetes from various locations |
 | |
| a: Condition factor; b: Growth coefficient; r2: Coefficient of determination | |
| Table 8: | Growth parameters (L ∝ and K) and computed growth performance index (φ/) of the genus Acetes from different tropical countries |
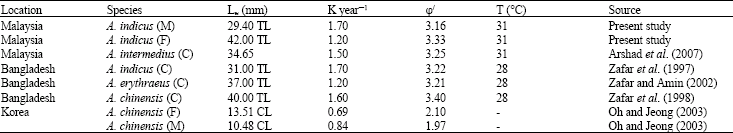 | |
| M: Male, F: Female, C: Combined sexes | |
The estimated ages from the study are presented in Table 6. The overall average growth rate of A. indicus showed 1.83 ± 0.78 mm/month for male and 2.30 ± 0.72 mm/month for females which enable it to attain a total length of around 21.96 mm for males and 27.6 mm for females in 12 months. Similar reports are available on other shrimp by Amin and Zafar (2003) and Cha et al. (2004) and on fish by Amin and Zafar (2004), Isaac and Ruffino (1996) and Blaber et al. (1998), who used length-converted age method for their study.
The recruitment pattern suggests that continuous recruitment consists of two peaks seasonal pulse (Fig. 9). This is in good agreement with Zafar et al. (1997) for A. indicus and Oh and Jeong (2003) for A. chinensis. There are no published reports on Acetes recruitment in Malaysia. However, it has been reported that the Acetes spawns throughout the year in the tropics and subtropics, spawning peaks can be recognized and these almost always lie in the warmer months (Nataraj, 1947). Spawning patterns in these areas (tropical and subtropical) are probably related to monsoonal influences on precipitation and wind direction (Omori, 1975). Major spawning was observed in February to March in the coastal waters of Malacca, Peninsular Malaysia (Fig. 6). It is expected that the major recruitment peak (September-October) detected in this study should correspond to the major spawning season.
On the basis of 175 independent data sets Pauly (1980) has made a regression analysis of M (year-1) on K (year-1), L ∝ and T (average annual temperatures). Sparre and Venema (1992) warned against using Pauly’s formula for crustaceans, mollusc or cephalopods or any other invertebrates, as the formula does not cover these groups. But in the absence of any data this formula can be used for shrimp (Niamaimandi et al., 2007). Pauly et al. (1984) noted that the formula can be used for shrimp and any other invertebrates, because these marine organisms generally share the same habitats, resources and predators and that therefore, they are not likely to differ widely in their vital parameters. Hence Pauly’s equation allows a rough estimation of M in shrimp stock where L ∝ , K and T are known.
The estimated natural mortality rate of A. indicus for both sexes in the present study (M = 2.65 year-1 in males and M = 1.91 year-1 in females) is higher than the estimated values of fishing mortality (F = 1.50 year-1in males and F = 1.59 year-1 in females). Higher natural mortalities of A. indicus verses the fishing mortalities indicate the unbalance position in the stock. This is based on the assumption that a stock is optimally exploited when fishing mortality (F) equals natural mortality (M), or E = (F/Z) = 0.5 (Gulland, 1971). Estimated exploitation level (E) of females (0.45) is higher than males (0.36) in A. indicus population. It is may be for the larger size of females they can not escape from the gear than males. In Bangladesh waters M and F have been estimated at 4.76 and 1.31 year-1 for the combined sexes of A. indicus (Zafar et al., 1997). Higher natural mortality than fishing mortality was also reported from other Acetes studies (Arshad et al., 2007; Zafar et al., 1998; Zafar and Amin, 2002; Oh and Jeong, 2003) in the Asian countries (Table 9). Hass et al. (2004) noted production mortality can be influenced by shrimp length, movement and the habitat type. Mortality due to predation is influenced by the characteristics of the predation population. Xu et al. (1995) reported that the influences of abiotic factors are probably the most important reasons affects of natural mortality.
In most short-lived species, it is difficult to identify the component age groups and estimate the numbers at each age. Therefore, for estimation of Z, length-frequency data are used and a length-frequency distribution can be converted to an age frequency distribution by means of a length-converted catch curve. Catch curve analysis is feasible, assuming that total mortality is constant from length upwards. This assumption could be violated by migration out of the fishing area of mature shrimp (Niamaimandi et al., 2007). In length-catch curve analysis for estimation of total mortality, the initial ascending data points are not included in the regression (Fig. 7a, b). These points represent younger age groups, which are subjected to a lower fishing mortality because they are either not fully recruited or not fully vulnerable to the fishing gear used (King, 1984).
| Table 9: | Mortality parameters (Z, F and M) and computed exploitation rate (E) of the genus Acetes from different tropical countries |
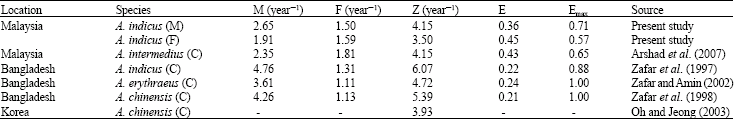 | |
| M: Male, F: Female, C: Combined sexes | |
The level of total mortality, Z = 6.07 year-1 from Bangladesh waters (Zafar et al., 1997) is higher than present study (Z = 4.15 year-1 for males and Z = 3.50 year-1 for females). The values of E = 0.36 for males and E = 0.45 for females are lower than optimum level of exploitation (E = 0.50), suggesting under-exploitation in this fishery (under-fishing occurs if M>F or E>0.50, Gulland, 1971).
Considering our relative Y/R and relative B/R analyses (Fig. 10a, b) our computed exploitation rate, E values (E = 0.36 for males and E = 0.45 for females) are below the predicted maximum values of Emax = 0.71 (males) and 0.57 (females). This reveals that the fishery is being under exploited in the coastal waters of Malacca. Results from the analysis of the exploitation rate (E) based on the fishing mortality estimates and from the relative Y/R, indicate that the fishery is below the optimum fishing pressure and probably the catch is below to the full exploitation (MSY). Thus, the more yield could be obtained by a reasonable increase in the effort. This implies that further increase in fishing effort could be possible to get the maximum sustainable yield, thus driving the stock sustain and economic benefit.
ACKNOWLEDGMENTS
This research is part of a Ph.D thesis funded by the Ministry of Science, Technology and Innovation (MOSTI), Malaysia (Grant No. 05-01-04-SF0613). The authors would like to thank to MOSTI for providing financial support to carry out this research work. Special thanks to Universiti Putra Malaysia for providing partial financial support through Graduate Research Fellowship (GRF) during the study period. In addition, thanks go to Mr. Ibrahim, fisherman and Mr. Perumal, Science Officer of Marine Science Laboratory and Aquaculture, Institute of Bioscience, UPM for the assistance during field sampling.
REFERENCES
- Nurul Amin, S.M., A. Arshad, J.S. Bujang and S.S. Siraj,, 2008. Growth, mortality and yield-per-recruit of sergestid shrimp, Acetes intermedius Omori, 1975 (Decapoda: Sergestidae) from length frequency analysis in the coastal waters of Malacca, Peninsular Malaysia. Pertanika J. Trop. Agric. Sci., 31: 55-66.
Direct Link - Arshad, A., S.M.N. Amin, S.S. Siraj and S.B. Japar, 2007. New distribution records of sergestid shrimp, Acetes intermedius (Decapoda: Sergestidae) from Peninsular Malaysia with notes on its population characteristics. J. Biol. Sci., 7: 1305-1313.
CrossRefDirect Link - Amin, S.M. and M. Zafar, 2004. Studies on age, growth and virtual population analysis of Coilia dussumieriI from the neritic water of Bangladesh. J. Biological Sci., 4: 342-344.
CrossRefDirect Link - Amin, S.M.N., M. Zafar and A. Halim, 2008. Age, growth, mortality and population structure of the oyster, Crassostrea madrasensis, in the Moheskhali Channel (Southeastern coast of Bangladesh). J. Applied Ichthyol., 24: 18-25.
CrossRefDirect Link - Botter-Carvalho, M.L., P.J.P. Santos and P.V.V.C. Carvalho, 2007. Population dynamics of Callichirus major (Say, 1818) (Crustacea: Thalassinidea) on a beach in northeastern Brazil. Est. Coast. Self Sci., 71: 508-516.
CrossRefDirect Link - Blaber, S.J.M., J. Staunton-Smith, D.A. Milton, G. Fry and T.V. Velde et al., 1998. The biology and life-history strategies of Ilisha (Teleostei: Pristigasteridae) in the coastal waters and estuaries of sarawak. Est. Coast. Shelf Sci., 47: 499-511.
CrossRefDirect Link - Cha, H.K., J.H. Choi and C.W. Oh, 2004. Reproductive biology and growth of the shiba shrimp, Metapenaeus joyneri (Decapoda: Penaeidae), on the western coast of Korea. J. Crust. Biol., 24: 93-100.
CrossRefDirect Link - Dall, W., B.J. Hill, P.C. Rothlisberg and D.J. Sharples, 1990. The biology of the Penaeidae. Adv. Mar. Biol., 27: 1-489.
Direct Link - Dulcic, J. and M. Kraljevic, 1995. Age, growth and mortality of damselfish (Chromis chromis L.) in the eastern middle Adriatic. Fish. Res., 22: 255-264.
Direct Link - Ecoutin, J.M., J.J. Albaret and S. Trape, 2005. Length-weight relationship for fish populations of a relatively undisturbed tropical estuary: The Gambia. Fish. Res., 72: 347-351.
CrossRefDirect Link - Enin, U.I., U. Lowenberg and T. Kunzel, 1996. Population dynamics of the estuarine prawn (Nematopalaemon hastatus Aurivillius 1898) off the southeast coast of Nigeria. Fish. Res., 26: 17-35.
CrossRef - Etim, L. and Y. Sankare, 1998. Growth and mortality, recruitment and yield of the fresh-water shrimp, Macrobrachium vollenhovenii, Herklots 1851 (Crustacea, Palaemonidae) in the Fahe reservoir, Cote d'Ivoire, West Africa. Fish. Res., 38: 211-223.
CrossRefDirect Link - Hass, H.L., A. Rose, B. Fry, J.J. Minello and L.P. Rozas, 2004. Brown shrimp on the edge: Linking habitat to survival using an individual based simulation model. Ecol. Appl., 14: 1232-1247.
Direct Link - Ikeda, T. and J.K.B. Raymont, 1989. Preliminary studies on the intermoult period and growth of the pelagic shrimp Acetes sibogae australis from a tropical sea. Bull. Plank. Soc. Jap., 36: 11-18.
Direct Link - Isaac, V.J. and M.L. Ruffino, 1996. Population dynamics of tambaqui, Colossoma macropomum Cuvier, in the Lower Amazon, Brazil. Fish. Manage. Ecol., 3: 315-333.
CrossRefDirect Link - Jayawardane, P.A.A.T., T.D.S. McLusky and P. Tytler, 2002. Estimation of population parameters and stock assessment of Penaeus indicus (H. Milne Edwards) in the Western coastal waters of Sri Lanka. Asian Fish. Sci., 15: 155-166.
Direct Link - Jayawardane, P.A.A.T., T.D.S. McLusky and P. Tytler, 2003. Population dynamics of Metapenaeus dobsoniI from the Western coastal waters of Sri Lanka. Fish. Manage. Ecol., 10: 179-189.
CrossRefDirect Link - King, J.E., 1984. A study of the reproductive organs of the common marine shrimp, Penaeus setiferus (Linnaeus). Biol. Bull., 94: 244-262.
PubMedDirect Link - Minello, T.J., R.J. Zimmerman and E.X. Martinez, 1989. Mortality of young brown shrimp, Penaeus aztecus in estuarine nurseries. Trans. Am. Fish. Soc., 118: 693-708.
CrossRefDirect Link - Moreau, J. and F.X. Cuende, 1991. On improving the resolution of the recruitment patterns of fishes. Fishbyte, 9: 45-46.
Direct Link - Munro, J.L. and D. Pauly, 1983. A simple method for comparing the growth of fishes and invertebrates. Fishbyte, 1: 5-6.
Direct Link - Niamaimandi, N., A.B. Arshad, S.K. Daud, R.C. Saed and B. Kiabi, 2007. Population dynamics of green tiger prawn, Penaeus semisulcatus (De Haan) in Bushehr coastal waters, Persian Gulf. Fish. Res., 86: 105-112.
CrossRefDirect Link - Oh, C.W. and I.J. Jeong, 2003. Reproduction and population dynamics of Acetes chinensis (decapoda: Sergestidae) on the Western coast of Korea, Yellow Sea. J. Crustacean Biol., 23: 827-835.
CrossRefDirect Link - Omori, M., 1978. Zooplankton fisheries of the world: A review. Mar. Biol., 48: 199-205.
CrossRefDirect Link - Paraconstantinou, C. and K. Kapiris, 2001. Distribution and population structure of the red shrimp (Aristeus antennatus) on an unexploited fishing ground in the Greek Ionian Sea. Aqua. Liv. Resour., 14: 303-312.
CrossRefDirect Link - Pauly, D., 1980. On the interrelationships between natural mortality, growth parameters and mean environmental temperature in 175 Fish stocks. J. Cons. Int. Explor. Mer., 39: 175-192.
CrossRefDirect Link - Pauly, D. and N. David, 1981. ELEFAN-I, a basic program for the objective extraction of growth parameters from Length frequency data. Meeresforschung, 28: 205-211.
Direct Link - Pauly, D. and J.L. Munro, 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte, 2: 21-21.
Direct Link - Smith, A.E., J.W. Chapman and B.R. Dumbauld, 2008. Population structure and energetics of the bopyrid isopod parasite Orthione griffennis in mud shrimp Upogebia pugettensis. J. Crust. Biol., 28: 228-233.
CrossRefDirect Link - Tsoumani, M., R. Liasko, P. Moutsaki, I. Kagalou and I. Leonardos, 2006. Length-weight relationships of an invasive cyprinid fish (Carassius gibelio) from 12 Greek lakes in relation to their trophic states. J. Applied Ichthyol., 22: 281-284.
CrossRefDirect Link - Uye, S.I., 1982. Length-weight relationships of important zooplankton from the Inland Sea of Japan. J. Ocean. Soc. Jpn., 38: 149-158.
CrossRefDirect Link