
H. Khazali
Medical School of Azad University, Tehran, Iran
Journal of Applied Sciences
Year: 2009 | Volume: 9 | Issue: 15 | Page No.: 2822-2828







ABSTRACT
The aim of this study was to determine whether galanin affects the mean plasma concentrations of metabolic parameters such as thyroxine (T4), triiodothyronine (T3), Growth Hormone (GH), insulin, in the young castrated camels fed different energy content in diets. Twenty four camels were randomly divided into 3 main groups. Animals of all groups were fed either 100, 50 or 25% energy content in diet for 30 days. After 30 days, camels were received daily single injections of either 1 or 2 ug galanin/kg b.wt. into their jugular veins for 5 days. Blood samples were collected daily from jugular veins of all animals before injections on day 4 until 4 days after the last injections of galanin. Samples were assayed for plasma T3, T4, GH and insulin concentrations by double-antibody RIA. Different energy contents in diet and injections of 1 and 2 ug galanin/kg b.wt. did not change the mean plasma concentrations of T3, T4 and urea in the animals of all groups. Twenty five percent energy content in diet for 30 days significantly (p<0.01) increased the mean plasma concentrations of GH and these increases were significantly (p<0.01) elevated by injection of 1 and 2 ug galanin/kg b.wt. Plasma levels of insulin was significantly (p<0.01) lower in those animals fed 25% energy and also injections of 1 and 2 ug galanin/kg b.wt. significantly (p<0.01) suppressed these low levels of insulin. The results of this experiment indicate that galanin may increase mean plasma concentrations of GH and decrease mean plasma concentrations of insulin in the camels as a pseudo-ruminants fed low energy content in diet that cause a severe body weight lost.
PDF Abstract XML References Citation
How to cite this article
H. Khazali, 2009. Effect of the Galanin on Growth Hormone, Thyroid Hormones and Insulin in Young Castrated Camelus dromedarius Fed Different Levels of their Energy Requirement. Journal of Applied Sciences, 9: 2822-2828.
DOI: 10.3923/jas.2009.2822.2828
URL: https://scialert.net/abstract/?doi=jas.2009.2822.2828
DOI: 10.3923/jas.2009.2822.2828
URL: https://scialert.net/abstract/?doi=jas.2009.2822.2828
INTRODUCTION
The desert camel (Camelus dromedarius), is a pseudo-ruminant, with certain physiological, biochemical and pharmacological peculiarities, which make them distinct from true ruminants. For example, higher plasma glucose level, less insulin, responsivity and fatty acid metabolism are some of the physiological peculiarities that differentiate them from ruminant (Harrison and Leat, 1975) Moreover, they are well known for their exceptional ability to withstand long periods of food deprivation However, the neural mechanisms enabling camels to withstand food deprivation are poorly understood. It is well established that orexigenic peptides secretions increase during energy deprivation in ruminants and nonruminants (Chaillou and Tillet, 2005). Among the orexigenic hormones, galanin is a 29 amino acid neuropeptide that is mostly found in ventral structures (Skofitsch and Jacobowitz, 1985; Melander et al., 1986). Based on its neuron distributions in ventral structures, galanin coexists with many other neurons. For instance, in hypothalamic area, galanin coexists with neurons secreting different neurotransmitters such as GHRH, GABA, noradrenaline, 5-hydroxytryptamine (5-HT) and NPY (Hokfelt et al., 1987; Holets et al., 1988). Therefore, galanin controls different physiological actions on many different tracts (Goldstein and Deutch, 1989; Tsuda et al., 1990; Kyrkouli et al., 1992). One of the physiological actions is its effect on metabolism and feeding behaviors that make galanin as an orexigenic hormone (Crawley et al., 1990; Corwin et al., 1993). The orexigenic effect of galanin decrease plasma levels of insulin, glucagon, somatostatin and gastrin and increase the release of growth hormone (Hermansen, 1988; Kaplan et al., 1988; Legakis, 2005; Merchentaler et al., 1990; Spencer et al., 1994; Stavroula et al., 2006). Most of the above studies were conducted in normal energy fed human and rat as a nonruminants and in sheep and goat as a ruminants. Pseudo-ruminants have different metabolism from that of nonruminants and ruminants (Harrison and Leat, 1975). It is also assumed that the control of feeding behavior is different in pseudo-ruminants from that of nonruminants and ruminants. There are very few reports about the orexigenic effect of galanin on the metabolic hormones in pseudo-ruminants fed different energy content in diet. Therefore, the first goal of this experiment was to determine whether galanin affects the mean concentrations of GH and insulin in the camels fed different energy content in diets. Among many studies done on the effect of galanin on metabolic hormone, there are no reports about the effect of the galanin on thyroid hormones. The importance of thyroid hormones in metabolism is well known. For example, thyroid hormones play an important role in the regulation of energy homeostasis via oxygen consumption and heat generation (De Jesus et al., 2001; Lanni et al., 2001). Changes in basal metabolic rate caused by different energy content in diet is accompanied by changes of thyroid hormones secretions. Therefore, the second goal of this study was to determine whether galanin alters the thyroid hormones secretion in the camels fed different energy content in diet.
MATERIALS AND METHODS
These experiments have been approved by the Committee of Research Ethics of Shahid Beheshti University. These experiments have also been at Shahid Beheshti University from 23rd of July 2006 untill 1st of July of 2007.
Animals and diets: Twenty four camels (weighing between 190-199 kg) were randomly divided into 3 main groups. Animals of all groups were fed either 100, 50 or 25% of their maintenance energy requirements for 30 days. All animals were fed every day at 09.00 o’clock, taking 1 to 3 h to finish up all of the days diets. After 30 days, camels were given daily single injections of either 1 or 2 ug galanin/kg b.wt. into their jugular veins for 5 days. Gross energy and chemical compositions of feedstuffs were analyzed in the Animal Science Research Institute of Yazd. Diets were formulated based on AFRC (1995) (Table 1). During the course of the experiment, daily feed was weighed based upon body weight and given individually to each camel every morning. The camels had free access to fresh water. Other requirements were balanced at maintenance level.
After 30 days, animals of all groups were anesthetized throughout the surgery for jugular vein cannulation. After surgery, on day 5, camels in groups 1, 3 and, 5 received 1 ug galanin and camels in groups 2, 4 and 6 received 2 ug galanin into their jugular vein for 5 days. The body weight of animals was measured on day 1 and 30 of the experiment.
| Table 1: | Experimental ration ingredients and content of energy and nutrients |
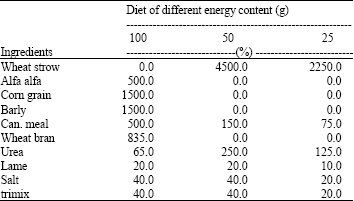 | |
Surgery for cannulation of the jugular vein: Surgical procedures were performed under general anesthesia induced by sodium pentobarbital and maintained by halothane in a closed circuit system (Dziuk et al., 1964). Catheters (70 cm) were made for the jugular vein with polysiloxane tubing with an inside diameter of 0.16 cm and a length of 70 cm. A piece of silk was attached to the catheter to secure the catheter to the tissue near the jugular vein. Camels were anesthetized with 30-40 mL of sodium pentobarbital (65 mg mL-1 concentration), scrubbed with iodized soap and sprayed with a tincture of prepodyne. The jugular vein was exposed as described by Pond and Houpt (1978). After cleaning the jugular vein, the catheter was inserted 15 cm into the vein. The silk was tied to tissue near the point of insertion. The catheter was passed subcutaneously to the dorsal aspect of the neck using a trocar. The incision was then closed with one stitch in the cut edge of the muscle, followed by suturing of the skin. A removable plastic plug was inserted into the external end of the catheter. No antibiotic were administered to the animals. Each camel was kept in a single cage for a 4 day recovery period. During recovery, catheters were flushed with PBS solution to prevent clotting.
Blood collection: Blood samples were collected from cannules that were put into the jugular veins, every day at 08:00 o’clock from 4 days before first injections of galanin until 4 days after the last galanin injections. Blood samples were kept at 4°C until centrifugation. A saturated sodium citrate solution (40 uL sodium citrate solution/ml blood) was added to the samples before centrifugation to prevent clotting of plasma during storage. Plasma was stored at -20°C until assayed for T3, T4, insulin and GH.
Hormone assays: Plasma T3, T4, insulin and GH were measured by a homologous double-antibody radioimmunoassay (RIA). An average binding percentage of 20 to 70% was performed for different hormone assays. The validity of all hormone assays was performed using a procedure earlier described by Kraeling et al. (1982) and Khazali (1992). For GH assay, bovine GH (TYN-BG) and antisera against GH were provided by Tabeshyarnoor Co., (Industrial City of Bu-Ali, Hamadan, Iran). Bovine GH (TYN-BG) was used for iodination. A 7-point standard curve ranging from 0.04 to 10 ng GH was used. An average assay binding of 20% was achieved using an initial 1:10000 dilution of GH antiserum for GH assays. The inter- and intra-assay variations were 7 and 8%, respectively. Insulin assay, bovine insulin (TYN-BI) and the antibody against insulin were provided by Tabeshyarnoor Co., (Industrial City of Bu-Ali, Hamadan, Iran). Bovine insulin (TYN-BI) was utilized for iodination. A 7-point standard curve ranging from 0.02 to 10 ng insulin was used. An average assay binding of 25% was achieved using an initial 1:2000 dilution of insulin antiserum for insulin assays. The inter- and intra-assay variations were 6 and 8%, respectively. For T3 assay, diiodothyronine was purchased from Sigma Chemical Company and T3 antisera was purchased from Chemicon Co., (Temmecula, Ca). T2 was used for iodination. A 6-point standard curve ranging from 0.32 to 5.2 ng T3/mL was used. An average assay binding of 70% was achieved with an initial 1:5000 dilution of T3 antiserum for T3 assays. The inter- and intra-assay variations were 7 and 8%, respectively. For T4 assay, T3 was purchased from Sigma Chemical Company and T4 antisera was purchased from Chemicon Co., (Temmecula, Ca). T3 was used for iodination. A 6-point standard curve ranging from 2.2 to 25 ng T4/mL was used. An average assay binding of 60% was achieved using an initial 1:5000 dilution of T4 antiserum for T4 assays. The inter- and intra-assay variations were 7 and 5%, respectively.
Statistical analysis: All analysis were conducted using General Linear Model procedures (SAS, 1996). Data was analyzed using an analysis of variance for a repeated measure design. Mean comparisons were evaluated by least significant difference with a single degree of freedom. A p<0.01 level of significance was chosen.
RESULTS
T3 and T4: Lower energy dietary intake and injections of 1 and 2 ug galanin/kg b.wt. did not change the mean plasma concentrations of the T3 and T4 in the animals of all groups (Fig. 1, 2). Mean plasma T3 and T4 levels in all animals were about 5.5 and 25 ng mL-1, respectively.
GH: Mean plasma GH levels of those camels fed 50 and 25% energy were 5 and 8.5 ng mL-1, respectively, that were significantly (p<0.01) higher than that of (3 ng mL-1) the animals fed 100% energy (Fig. 3).
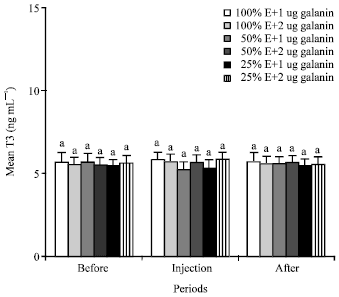 | |
| Fig. 1: | Mean plasma concentrations of T3 of the animals in the different groups of 1 (NE and 1 ug galanin), 2 (NE and 2 ug galanin), 3 (LE and 1 ug galanin) and 4 (LE and 1 ug galanin) and before, during and after infusions of galanin. Treatments with different letter(s) are significantly different at p<0.01, E: Energy |
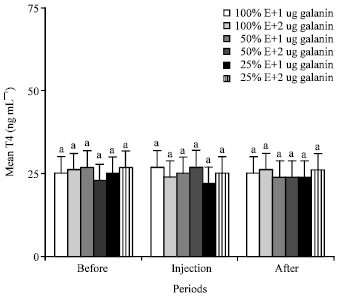 | |
| Fig. 2: | Mean plasma concentrations of T4 of the animals in the different groups of 1 (NE and 1 ug galanin), 2 (NE and 2 ug galanin), 3 (LE and 1 ug galanin) and 4 (LE and 1 ug galanin) and before, during and after infusions of galanin. Treatments with different letter(s) are significantly different at p<0.01, E: Energy |
Injections of 1 and 2 ug galanin/kg b.wt. significantly (p<0.01 ) increased the mean plasma GH level from 8.5 to 13 and 15 ng mL-1 in the animals fed 25% energy, but not in those animals fed 100% and 50% energy (Fig. 3).
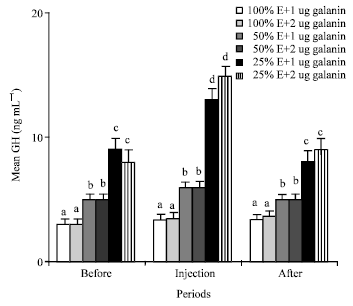 | |
| Fig. 3: | Mean plasma concentrations of GH of the animals in the different groups of 1 (NE and 1 ug galanin), 2 (NE and 2 ug galanin), 3 (LE and 1 ug galanin) and 4 (LE and 1 ug galanin) and before, during and after infusions of galanin, a, b, c and d: Treatments with different letter(s) are significantly different at p<0.01, E: Energy |
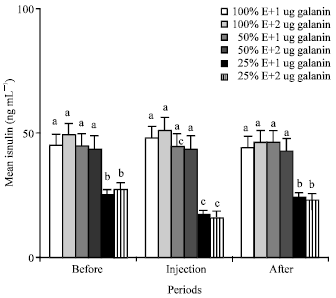 | |
| Fig. 4: | Mean plasma concentrations of insulin of the animals in the different groups of 1 (NE and 1 ug galanin), 2 (NE and 2 ug galanin), 3 (LE and 1 ug galanin) and 4 (LE and 1 ug galanin) and before, during and after infusions of galanin. a, b and c: Treatments with different letter(s) are significantly different at p<0.01, E: Energy |
Insulin: Mean plasma concentrations of the insulin of the camels fed 50% energy were similar to those camels fed 100% energy of their requirement (Fig. 4). Also, injections of 1 and 2 ug galanin/kg b.wt. did not change the mean plasma levels in the animals fed 100 and 50% energy (Fig. 4). Twenty five percent energy content in diet and injections of 1 and 2 ug galanin/kg b.wt. significantly (p<0.01) decreased mean plasma concentrations of insulin from 45 to 25 and from 25 to 16 ng mL-1 in those animals of groups 5 and 6 (Fig. 4).
DISCUSSION
Lower energy dietary intake and injections of 1 and 2 ug galanin/kg b.wt. did not change the mean plasma concentrations of the T3 and T4 in the animals of all groups (Fig. 1, 2). Present study is the first to report the effect of galanin on thyroid hormones in the pseudo-ruminants. The results of the effect of galanin on mean plasma T3 and T4 levels of the camels fed different energy content in diet are different from the results of earlier findings that reported lower energy dietary intake decreased the plasma thyroid hormones of nonruminants (Khazali, 1992) and ruminant (Takasu et al., 2007). Also, the results in present study of the effect of galanin on thyroid hormone concentrations are different from those of the study of Legradi et al. (1997), which reported the peripheral injection of galanin increased the plasma level of Thyroid Stimulating Hormones (TSH) in nonruminants such as rat and human, but there was no data on the plasma level of T3 and T4 in that study. It is well established that the increase of plasma TSH level is accompanied by the decrease of plasma T3 and T4 in 100% energy-fed human (Felig and Frohman, 2001; Reasner and Ralbert, 2002).
In nonruminants and ruminants, the Hypothalamus Pituitary Thyroid (HPT) axis plays an important role in the regulation of energy homeostasis (De Jesus et al., 2001; Lanni et al., 2001) via the effects of the thyroid hormone to increase oxygen consumption and heat generation (De Jesus et al., 2001; Lanni et al., 2001). Thus inhibition of the HPT axis during fasting would appear to be an important adaptive mechanism to conserve energy stores (Rondeel et al., 1992; Van Haasteren et al., 1995; Legradi et al., 1997). The state of central hypothyroidism induced by fasting is orchestrated by changes of circulating levels of galanin, which rise with fasting and is restored to normal levels by refeeding (Rondeel et al., 1992). Thus, if galanin is administered exogenously to fasting animals, the more decrease in circulating levels of thyroid hormones can be observed (Legradi et al., 1998).
Present results indicate that the 100, 50 and 25% energy-fed camels as pseudo-ruminants are not as sensitive to galanin as those as nonruminants and ruminants. Even when the camels are in a long term fasting period, they are not sensitive to the effect of galanin injections. Therefore, we should consider that galanin has no physiological actions in thyroid metabolism of long-term fasted pseudo-ruminants.
GH: Mean plasma GH levels of those camels fed 50 and 25% energy were significantly (p<0.01 ) higher than that of (3 ng mL-1) the animals fed 100% energy (Fig. 3). These results are similar to the previous finding (Moravej et al., 2006) that indicated lower energy content in diet increase GH level. This may be due to effect of GH on adipose tissue breakdown to release fatty acid as a metabolic fuel. Injections of 1 and 2 ug galanin/kg b.wt. significantly (p<0.01) increased the mean plasma GH level in the animals fed 25% energy (Fig. 3). Present study is the first to report the effect of galanin injections on GH in the pseudo-ruminants fed different energy content in diets. Present result about the effect of galanin on GH in the camels fed 25% energy is similar to other studies indicating that galanin is a hypophysiotropic hormone that elicits GH secretion (Bauer et al., 1986; Giustina et al., 1993) and enhances the GH response to GHRH in 100% energy fed nonruminant (Davis et al., 1987). Furthermore, conflicting evidences exist about the direct effect of galanin on GH, with an inhibitory influence on GH secretion observed in rat (Meister and Hulting, 1987) and a stimulatory one observed in rat (Lindstromm and Sävendhal, 1993) and bovine (Baratta et al., 1997). Present findings about the effect of galanin on GH in the camels fed 100 and 50% energy are different compared to the results of other studies that showed injections of galanin increase GH in rat and human (Kaplan et al., 1988; Merchentaler et al., 1990). This may be due to normal plasma level of insulin and the inhibitory effect of normal concentrations of plasma glucose on the GH secretions (Holl et al., 1999) in the camels fed 100 and 50% energy.Insulin: Mean plasma concentrations of the insulin of the camels fed 50% energy were similar to those camels fed 100% energy of their requirement (Fig. 4). These results are different from those earlier findings (McDonald et al., 1985) that indicated lower energy content in diet decreased insulin levels in ruminants. This may be due to the normal plasma glucose concentrations of the animals. Also, injections of 1 and 2 ug galanin/kg b.wt. did not change the mean plasma levels in the animals fed 100 and 50% energy (Fig. 4). Earlier findings also reported that galanin may slightly decrease the plasma level of insulin in nonruminant (Stavroula et al., 2006). In those studies, the effect of galanin was not on the long term fasting subject that we had in present experiment. Twenty five percent energy content in diet and injections of 1 and 2 ug galanin/kg b.wt. significantly (p<0.01) decreased mean plasma concentrations of insulin from 45 to 25 and from 25 to 16 ng mL-1 in those animals of group 5 and 6 (Fig. 4). Present results are similar to the previous finding reporting that Intravenous administration of galanin into fasted conscious dogs as a nonruminant, decreased plasma insulin levels (McDonald et al., 1985). The mechanism of inhibitory effect of galanin on insulin release most likely occurs through the inhibition of adenylate cyclase, involving a petussis-toxin-sensitive inhibitory GTP-binding regulatory protein and the activity of protein kinase C and cyclic AMP (Amiranoff et al., 1988; Lindskog and Ahren, 1991; Marsoobian et al., 1995; Leibowitz et al., 1998).
CONCLUSION
The results of this study indicated that in contrast to the effects of galanin on metabolic parameters in ruminants and nonruminants, galanin has no physiological effects on metabolic parameters in normally fed camels. This may be due to the certain physiological, biochemical and pharmacological peculiarities such as their extraordinary ability to undergo long periods of food deprivation that make them distinct from other related ruminants and nonruminants. Galanin may increase the plasma levels of GH and decrease the plasma levels of insulin in the camels as pseudo-ruminants, but only when they are in severe negative energy balance leading to severe body weight loss. On the contrary, ruminants and nonruminants are responsive to the galanin even when their metabolic rates are in positive energy balance.
REFERENCES
- Chaillou, E. and Y. Tillet, 2005. Nutrition and hypothalamic neuropeptides in sheep: Histochemical studies. Histol. Histopathol., 20: 1209-1225.
Direct Link - De Jesus, L., S. Carvalho, M. Ribeiro, M. Schneider, S. Kim, J. Harney and P. Bianco, 2001. The type 2 iodothyronine deiodinase is essential for adaptive thermogenesis in brown adipose tissue. J. Clin. Invest., 108: 1379-1385.
Direct Link - Goldstein, M. and A. Deutch, 1989. The inhibitory actions of NPY and galanin on 3H-norepinephrine release in the central nervous system: Relation to a proposed hierarchy of neuronal coexistence. Proceedings of the Karolinska Institute Nobel Conference Series, September 7-8, 1989, Stockholm, Sweden, pp: 153-162.
- Harrison, F.A. and W.M. Leat, 1975. Digestion and absorption of lipid in non-ruminant and ruminant animals: A comparison. Proc. Nutr. Soc., 34: 203-210.
PubMed - Lanni, A., M. Moreno, A. Lombardi, P. de Lange and F. Goglia, 2001. Control of energy metabolism by iodothyronines. J. Endocrinol. Invest., 24: 897-913.
Direct Link - Legakis, I., 2005. The role of Galanin in metabolic disorders leading to type 2. Diabetes Mellitus Drug News Perspect., 18: 173-178.
Direct Link - Legradi, G., C.H. Emerson, R.S. Ahima, J.S. Flier and R.M. Lechan, 1997. Leptin prevents fasting-induced suppression of prothyrotropin-releasing hormone messenger ribonucleic acid in neurons of the hypothalamic paraventricular nucleus. Endocrinology, 138: 2569-2576.
CrossRefPubMedDirect Link - Legradi, G., C.H. Emerson, R.S. Ahima, W.M. Rand, J.S. Flier and R.M. Lechan, 1998. Arcuate nucleus ablation prevents fasting-induced suppression of ProTRH mRNA in the hypothalamic paraventricular nucleus. Neuroendocrinology, 68: 89-97.
Direct Link - Moravej, H., H. Khazali, M. Shivazad and H. Mehrabani-Yeganeh, 2006. Plasma concentrations of thyroid hormone and growth hormone in lohmann male broilers fed on different dietary energy and protein levels. Int. J. Poult. Sci., 5: 457-462.
CrossRefDirect Link - Stavroula, E., H. Kyrkouli, H. Strubbe and A. Scheurink, 2006. Galanin in the PVN increases nutrient intake and changes peripheral hormone levels in the rat. Physiol. Behav., 89: 103-109.
Direct Link - Takasu, M., Y. Kishima, N. Nishii, Y. Ohba, S. Maeda, K. Miyazawa, T. Murase and H. Kitagawa, 2007. Insulin and glucagon secretory patterns during propionate and arginine tolerance tests in Japanese black cattle with growth retardation. Vet. Med. Sci., 69: 69-72.
Direct Link - Van Haasteren, G., E. Linkels, W. Klootwijk, H. van Toor and J. Rondeel et al., 1995. Starvation induced changes in the hypothalamic content of prothyrotrophin-releasing hormone (proTRH) mRNA and the hypothalamic release of proTRH-derived peptides: Role of the adrenal gland. J. Endocrinol., 145: 143-153.