
Taghi Naserpour Farivar
Department of Microbiology, Zahedan University of Medical Sciences, Zahedan, Iran
Batool Sharifi Moud
Department of Infectious Disease, School of Medicine, Research Institute of Infectious Diseases and Tropical Medicine, Zahedan University of Medical Sciences, Zahedan, Iran
Mansur Sargazi
Department of Clinical Biochemistry and Metabolic Medicine, Royal Liverpool University Hospital, Liverpool, UK
Alireza Moeenrezakhanlou
Division of Infectious Diseases, Department of Medicine, Vancouver Coastal Health Research Institute, University of British Columbia, Vancouver, British Columbia, Canada
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 4 | Page No.: 719-722







ABSTRACT
In this study, we aimed to determine the significance of association between Tuberculosis and apolipoprotein E polymorphism. The apolipoprotein E genotypes were assayed in 250 tuberculosis patients by polymerase chain reaction followed by enzymatic digestion with Hha I. The results were compared with the results of the same experiments on 250 sex and age matched control peoples. Present results showed that in studied populations, prevalence of E4 genotype was lower in controls than in patients (8 v. 13.2%; OR = 1.75, p<0.05) and prevalence of E3 genotype was high in controls than in patients (86 v.51%; OR = 0.17, p<0.05). Statistically significant difference was found between patients and controls with respect to ε2 allele frequencies, while ε2 allele frequency was found to be much less prevalent in controls (6%) than in patients (35.8%; OR = 8.72, p<0.05). Also, our study revealed that there is an association between apolipoprotein E genotypes and amplitude to tuberculosis in studied populations. However, large population-based studies are needed to understand the exact role played by the locus in causing the condition.
PDF Abstract XML References Citation
How to cite this article
Taghi Naserpour Farivar, Batool Sharifi Moud, Mansur Sargazi and Alireza Moeenrezakhanlou, 2008. Apolipoprotein E Polymorphism in Tuberculosis Patients. Journal of Applied Sciences, 8: 719-722.
DOI: 10.3923/jas.2008.719.722
URL: https://scialert.net/abstract/?doi=jas.2008.719.722
DOI: 10.3923/jas.2008.719.722
URL: https://scialert.net/abstract/?doi=jas.2008.719.722
INTRODUCTION
Apolipoprotein E (Apo E) is an exchangeable protein which acts as ligand for Low-Density Lipoprotein (LDL) receptors. It also has a repair function in response to tissue injury. ApoE plays an essential role in lipid metabolism, especially in the removal of atherogenic remnants of triglyceride-rich lipoproteins and by reversing cholesterol transport in plasma and intercellular lipid transport within tissues (Siest et al., 1995).
The human Apo E gene is 3.7 kb including 4 exons and 3 introns (Paik et al., 1985) Apolipoprotein E which is encoded by distinct APOE ε2, ε3 and ε4 has three major isoforms and these isoforms differ with each other in amino acid sequence at their 112 and 158. Each ApoE genotype has a specific amino acid characteristics. ApoE2 has two Cysteine, ApoE4 two arginine and ApoE3 one Cysteine and one arginine at their two position. Also, it has been reported that there is a direct relation between gene polymorphism and concentration of apoE (Zetterberg et al., 2004; Blennow et al., 2000).
The lymphocyte modulatory action of ApoE was originally discovered as an immune-inhibitory activity of LDL in vitro (Paik et al., 1985). Curtiss and his colleagues have shown that ApoE inhibits both luteinizing hormone-stimulated androgen production by ovarian theca cells and the proliferation of activated lymphocytes (Dyer and Curtiss, 1988). Also, it has been shown that ApoE is produced by macrophages (Hansson et al., 1989), which activate T lymphocytes by antigen presentation, while activated T cells secrete interferon-γ (IFN- γ), which in turn inhibits macrophage ApoE expression (Mistry et al., 1995). This implies an intricate, ApoE-mediated feedback regulation of immune activation. On the hand, our present information about ApoE and it`s role as an immune modulator is based on cell culture experiments and little is known about its function in integrative immune regulation (Hansson et al., 1989; Natasja de Bont et al., 1999).
Although it has been known for many years that ApoE can modulate immune activation, its immunological mechanism of action has remained unclear (Tenger and Xinghua, 2003). It has been showed that ApoE deficiency results in enhanced expression of immunostimulatory cell surface molecules by the activated APC. The expression of MHC molecules that present T-cell antigens, together with the co-stimulatory surface proteins CD40 and CD80 are crucial for activation of the T-cells. By modulating the level of these molecules, ApoE may time-tune the T-cell response to antigens (Tenger and Xinghua, 2003).
On the other hand, previous studies has been showed that IFN-γ secretion further activates APC to express more MHC class II and co-stimulatory molecules and therefore enhanced APC-T-cell interaction. The lack of ApoE caused stronger proliferation of the T cells and higher secretion of IFN-γ in vitro and increased expression of MHC class II and co-stimulatory molecules after IFN-γ stimulation in vivo. It is known that macrophages can produce ApoE and also have receptors that can recognize ApoE protein (Kelly et al., 1994). The ApoE secreted by macrophages may therefore dampen immune activation by inhibiting the expression of MHC and co-stimulatory molecules via an autocrine loop (Kelly et al., 1994).
On the other side, it has been showed that Mycobacterium tuberculosis which is an intracellular pathogen, persists in phagocyte cells by inhibiting maturation of the phagosome. Also, Gutierrez et al. (2004) showed that autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. His study indicated that the induction of autophagy (an intracellular pathway that facilitates the turnover of cytoplasmic macromolecules and organelles) can overcome the blockage in phagosomal maturation and that the antimycobacterial effects of interferon-γ (IFN-γ) can, in part, be explained by its ability to stimulate autophagy (Gutierrez et al., 2004).
In addition to its effects on T-cell activation, ApoE modulates the innate immune response in mice. As shown by Roselaar (Natasja de Bont et al., 1999) and De Bont (Simon et al., 1998) and their colleagues, E0 mice are more sensitive to infection with the intracellular bacterium, Listeria monocytogenes and Klebsiella pneumoniae, suggesting that ApoE promotes the macrophage-dependent defense against micro-organisms. ApoE may therefore exert the paradoxical effects of enhancing innate and dampening adaptive immune responses. Such effects could perhaps be explained by ApoE-dependent control of the differentiation and/or activation of macrophages. As association of apolipoprotein E genotypes with different infectious diseases has been showed before (Price et al., 2006; Chi et al., 2004).
So, in this study we aimed to determine the significance of association between susceptibility to Tuberculosis and apolipoprotein E genotypes in patients and control individuals.
MATERIALS AND METHODS
The apolipoprotein E genotypes were assayed in 250 patients (with out respect to their stage of disease and history of previous infection) and compared with 250 controls by polymerase chain reaction followed by enzymatic digestion with Hha I. Using chai test and odds ratio.
Apo E genotyping: Leukocyte DNA was extracted following standard protocols (Miller et al., 1988). DNA was amplified by PCR in a DNA thermal cycler (Eppendorf) using oligonucleotide primers (DNA Technology A/S,Denmark), forward (5`-ACAGAATTCGCCCCGGCCTGGTACAC-3`) and reverse (5`-TAAGCTTGGCACGGCTGTCCAAGGA-3`) as described by Hixson and Vernier (1990). Electrophoresis of amplified products (244 bp) was performed on 14% polyacrylamide gel, after digesting with the Hha I restriction enzyme (10 μL PCR product + 5 units of enzyme incubated at 37°C overnight). After electrophoresis, the gel was stained with ethidium bromide (0.2 mg L-1) for 10 min and the digested fragments were visualized under UV illumination. Sizes of Hha1 fragments (91, 83, 72, 48, 38 and 35 bp) were determined by comparison with known size marker (50 bp).
Statistical analysis: Standard statistical procedures from the SPSS package were used for the analysis of the data. Allele frequencies were calculated as per gene frequency formula and following Hardy-Weinberg law and the frequencies were tested for Hardy-Weinberg equilibrium. The allelic association was further tested by Wolf `s test. Multivariate adjusted odds ratios were calculated for apo ε4 allele (ε4/4 and ε3/4) and ε2 allele (ε2/2, ε2/3 and ε2/4) taking ε3 allele (ε3/3) as reference. For each odds ratio we calculated two tailed p-values and 95% Confidence Intervals (CI). Significance levels were set at 0.05 in all cases.
RESULTS AND DISCUSSION
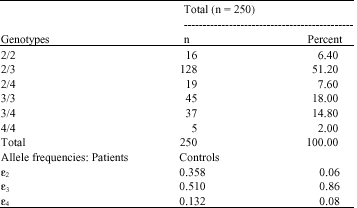
All PCR amplifications showed an expected fragment size (244 bp) on agarose gel electro-phoresis. Each genotype was determined by the specific combination of fragments generated from restriction endonuclease cut. Except for a 38 bp common fragment, other 16 and 18 bp common fragments were too small to detect. The codon changes from cysteine residues (GTCG) to arginine residues (GCGC) makes HhaI cleavage sites at amino residues position 112 and 158 on apo E alleles. The E2 isoform has cysteine at residues 112 and 158, so the ε2 allele had only two big fragments (91 and 83 bp). Whereas due to the substitution from arginine to cysteine residues at position 158 in E3, the 83 bp fragment was digested to 48 and 35 bp fragments in the ε3 allele. In addition to ε3, E4 has arginine at position 112; therefore, the 91 bp fragment was digested to a 72 bp fragment in the ε4 allele (the 19 bp fragment was too small for detection). Therefore, each genotype generates unique combinations of HhaI fragment sizes. The E2/E2 sample contained 91 and 83 bp HhaI fragments reflecting the absence of sites at 112 Cys and 158 Cys. The E3/E3 sample also contained the 91 bp fragment (112 Cys), as well as 48 and 35 bp fragments from HhaI cleavage at 158 Arg site. The E4/E4 sample also contained 48 and 35 bp fragments (158 Arg), as well as a unique 72 bp fragment from cleavage at 112 Arg (The 19 bp fragment was too small for detection). Heterozygotic alleles contained both sets of fragments from each apo E allele (Hixson and Vernier, 1990). Distribution of apo E genotypes and allele frequencies are shown in Table 1. All control`s apo E genotypes and allele frequencies in the present study were similar to those of other previous reported Iranian (Eichner et al., 2002) and some other of Asian (Gerdes et al., 1992) reported study.
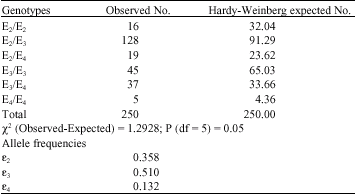
It was observed that prevalence of E4 genotypes was lower in controls when compared to patients (8 v. 13.2%; OR = 1.75, p<0.05) and prevalence of E3 genotypes was high in controls than in patients (86 v.51%; OR = 0.17, p<0.05) (Table 1). Statistically significant difference was found between patients and controls with respect to ε2 allele frequencies, while ε2 allele frequency was found to be much less prevalent in controls (6%) than in patients (35.8%; OR = 8.74, p<0.05). Further, the allelic distribution was found to be deviated significantly from Hardy-Weinberg equilibrium in patients (Table 2).
Studies conducted in different parts of the globe reveal that the gene frequencies at apo E locus are highly heterogeneous between the populations. The ε3 is the most common form of the gene in most of the populations followed by ε4 and ε2 alleles (Hallman et al., 1991). Also, different studies showed different results in the role of ApoE and it`s genotypes on susceptibility and resistance to different infectious disease. Hill et al. (2007) suggested that functional APOE gene polymorphisms may be a determinant of outcome in HCV infection and they hypothesized that the ε2 allele may protect against viral persistence via defective binding of HCV lipoviral particles to the cellular receptors involved in entry of these infectious particles. Although precise mechanism of ApoE effects in Mycobacterial interaction with Immunity system is not cleared in detail, bur Davis et al. (2002) showed that Mycobacterium marinum infection redirects the developmental program of embryonic macrophage migration. These infected macrophages, undergo a distinct phenotypic transformation into early microglial cells that express high levels of Apolipoprotein E. Also, Huang et al. (2004) suggested a protective effect (antioxidative effect) for ApoE3 and a detrimental effect (Oxidative stress) for ApoE4 to HSV-1 infection. On the other hand, previous studies (Natasja de Bont et al., 1999; Simon et al., 1998; Tarr et al., 2005) addressed a role, although indirect, for ApoE and it`s genotypes on susceptibility and resistance to different infectious disease. Also, Gerard et al. (1999) study showed that individuals bearing at least one copy of the APOE ε4 allele may be at increased risk for synovial infection by C. pneumoniae.
| Table 1: | Prevalence of ApoE genotypes and allelic frequencies in tuberculosis patients |
 | |
| Table 2: | Distribution of the Apo E genotypes and allelic frequency in tuberculosis patients |
 | |
CONCLUSION
In accordance with these findings, our results showed that in tuberculosis patient allelic frequencies of ApoE2 and ApoE4 were more prevalent than controls and ApoE3 genotype was more prevalent in controls than in patients, but much more studies in different population must been done to verify our results.
REFERENCES
- Blennow, K., A. Ricksten and J.A. Prince et al., 2000. No association between the 2-macroglobulin (2M) deletion and Alzheimer’s disease and no change in 2M mRNA, protein, or protein expressions. J. Neural Transm., 107: 1065-1079.
Direct Link - Chi, H., E. Messas, R.A. Levine, D.T. Graves and S. Amar, 2004. Interleukin-1 receptor signaling mediates atherosclerosis associated with bacterial exposure and/or a high-fat diet in a murine apolipoprotein E heterozygote model: Pharmacotherapeutic implications. Circulation, 110: 1678-1685.
Direct Link - Davis, J.M., H. Clay, J.L. Lewis, N. Ghori, P. Herbomel and L. Ramakrishnan, 2002. Real-Time visualization of Mycobacterium-Macrophage interactions leading to initiation of granuloma formation in Zebrafish embryos. Immunity, 17: 693-702.
Direct Link - Dyer, C.A. and L.K. Curtiss, 1988. Apoprotein E-rich high density lipoproteins inhibit ovarian androgen synthesis. J. Biol. Chem., 263: 10965-10973.
PubMedDirect Link - Eichner, J.E., S.T. Dunn, G. Perveen, D. M. Thompson, K.E. Stewart and B.C. Stroehla, 2002. Apolipoprotein E polymorphism and cardiovascular disease: A HuGE review. Am. J. Epidemiol., 155: 487-495.
CrossRefDirect Link - Gutierrez, M.G., S.S. Master, S.B. Sing G. Taylor, M. Colombo and V. Deretic, 2004. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell, 119: 753-766.
Direct Link - Hallman, D.M., E. Boerwinkle, N. Saha, C. Sandholzer, H.J. Menzel, A. Csazar and G. Utermann, 1991. The apolipoprotein E polymorphism: A comparison of allele frequencies and effects in nine populations. Am. J. Hum. Genet., 49: 338-349.
PubMedDirect Link - Hansson, G.K., J. Holm and L. Jonasson, 1989. Detection of activated T lymphocytes in the human atherosclerotic plaque. Am. J. Pathol., 135: 169-175.
PubMed - Hill, J.M., P.S. Bhattacharjee and D.M. Neumann, 2007. Apolipoprotein E alleles can contribute to the pathogenesis of numerous clinical conditions including HSV-1 corneal disease. Exp. Eye Res., 84: 801-811.
Direct Link - Hixson, J.E. and D.T. Vernier, 1990. Restriction isotyping of human apolipoprotein E by gene amplification and cleavage with HhaI. J. Lipid Res., 31: 545-548.
PubMedDirect Link - Huang, Y., K.H. Weisgraber, L. Mucke and W. Robert Mahley, 2004. Apolipoprotein E: Diversity of cellular origins, structural and biophysical properties and effects in Alzheimer's disease. J. Mol. Neurosci., 23: 189-204.
Direct Link - Miller, S.A., D.D. Dykes and H.F. Polesky, 1988. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res., 16: 1215-1215.
PubMedDirect Link - Mistry, M.J., M.A. Clay, M.E. Kelly, M.A. Steiner and J.A. Harmony, 1995. Apolipoprotein E restricts interleukin-dependent T lymphocyte proliferation at the G1A/G1B boundary. Cell Immunol., 160: 14-23.
PubMed - de Bont, N., M.G. Netea, P.N. Demacker, I. Verschueren and B.J. Kullberg et al., 1999. Apolipoprotein E knock-out mice are highly susceptible to endotoxemia and Klebsiella pneumoniae infection. J. Lipid Res., 40: 680-685.
PubMedDirect Link - Price, D.A., M.F. Bassendine, S.M. Norris, C. Golding and G.L. Toms et al., 2006. Apolipoprotein ε3 allele is associated with persistent hepatitis C virus infection. Gut, 55: 715-718.
CrossRefDirect Link - Siest, G., T. Pillot, A. Regis-Bailly, B. Leininger-Muller, J. Steinmetz, M.M. Galteau and S. Visvikis, 1995. Apolipoprotein E: An important gene and protein to follow in laboratory medicine. Clin. Chem., 41: 1068-1086.
PubMed - Roselaar, S.E. and A. Daugherty, 1998. Apolipoprotein E-deficient mice have impaired innate immune responses to Listeria monocytogenes in vivo. J. Lipid Res., 39: 1740-1743.
PubMedDirect Link - Tarr, P.E., P. Taffe, G. Bleiber, F. Hansjakob and R. Margalida et al., 2005. Modeling the influence of APOC3, APOE and TNF polymorphisms on the risk of antiretroviral therapy-associated lipid disorders. J. Infect. Dis., 191: 1419-1426.
Direct Link - Tenger, C. and Z. Xinghua, 2003. Apolipoprotein E modulates immune activation by acting on the antigen-presenting cell. Immunology, 109: 392-397.
Direct Link - Zetterberg, M., H. Zetterberg, M. Palmer, L. Rymo and K. Blennow et al., 2004. Apolipoprotein E polymorphism in patients with cataract. Br. J. Ophthalmol., 88: 716-718.
CrossRef