
O.B. Akpor
Department of Environmental, Water and Earth Sciences, Tshwane University of Technology,P/Bag X680, Pretoria, 0001, South Africa
M.N.B. Momba
Department of Environmental, Water and Earth Sciences, Tshwane University of Technology,P/Bag X680, Pretoria, 0001, South Africa
J. Okonkwo
Department of Environmental, Water and Earth Sciences, Tshwane University of Technology,P/Bag X680, Pretoria, 0001, South Africa
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 489-495







ABSTRACT
This study was aimed at investigating the effect of nine different nutrient/carbon supplements in mixed liquor on nutrient uptake ability of three wastewater protozoan isolates, which have previously been screened for phosphate and nitrate uptake efficiency. The results revealed that over 50% of phosphate was removed in the presence of sodium acetate, glucose or sucrose. Similarly, nitrate uptake of over 60% was observed in the presence of sodium acetate, sodium succinate, glucose or sucrose. These trends were common in all the isolates. Chemical Oxygen Demand (COD) removal in the mixed liquor was only found to be significantly removed in mixed liquors that were supplemented with glucose, sucrose or sodium succinate. In the presence of sodium acetate, COD was observed to increase. The findings of this investigation have revealed that nutrient uptake and COD removal by the test protozoan isolates may be dependent primarily on the initial nutrient supplement in mixed liquor.
PDF Abstract XML References Citation
How to cite this article
O.B. Akpor, M.N.B. Momba and J. Okonkwo, 2008. Effect of Nutrient/Carbon Supplements on Biological Phosphate and Nitrate Uptake by Protozoan Isolates. Journal of Applied Sciences, 8: 489-495.
DOI: 10.3923/jas.2008.489.495
URL: https://scialert.net/abstract/?doi=jas.2008.489.495
DOI: 10.3923/jas.2008.489.495
URL: https://scialert.net/abstract/?doi=jas.2008.489.495
INTRODUCTION
Wastewater often contains large amounts of nutrients, such as nitrogen and phosphorus in the form of nitrate and phosphate, which promote plant growth. Water that gets contaminated with nitrates can prove fatal, especially to infants that drink formula milk, as it restricts the amount of oxygen that reaches the brain, causing the blue baby syndrome. The presence of nitrate in water also causes digestive tract ulcers and algae bloom, resulting in eutrophication of surface waters (Kross et al., 1993). The presence of phosphate in water has a major impact on the environment (Foy and Withers, 1995).
Biological nutrient removal is known to be an effective and economical way to remove phosphorus, along with nitrogen and organic materials from wastewater (Grady et al., 1999). The roles protozoa play in nutrient removal in aquatic systems have only evolved in recent years. Initially, bacteria were considered the primary agents for the mineralization of major nutrients, but this concept has changed and it is now known that a significant fraction of nutrient mineralization taking place in nutrient removal systems may be a result of protozoan activity (Coleman, 1994; Caron et al., 1998).
The presence of an appropriate carbon source is known to affect nutrient removal from wastewater. Although a number of studies (Sang et al., 1997; Kargi et al., 2005) have reported the effects of carbon sources on nutrient removal performance, but not much is known on the effect of carbon/nutrient sources on phosphate and nitrate removal by protozoa. This study was aimed at investigating the following:
| • | Determining the effect of several carbon/nutrient supplements in mixed liquor on phosphate and nitrate uptake efficiency of three protozoan isolates. |
| • | Establishing the role of the protozoa in Chemical Oxygen Demand (COD) and Dissolved Oxygen (DO) concentration of the mixed liquor, in the presence of the carbon/nutrient supplements. |
MATERIALS AND METHODS
Test protozoan isolates: Three protozoan isolates were used in this investigation. The isolates were obtained from the aerobic zone of Daasport wastewater treatment plant in Pretoria South, in November 2006 and have previously been screened for nutrient removal efficiency (Akpor et al., 2007). The isolates were two ciliates and one flagellate and were presumptively identified as Chilophyllum sp., Aspidisca sp. and Paranema sp., respectively. In this study, isolates have been referred to as A, B and C, to represent Chilophylum sp., Aspidisca sp. and Paranema sp., respectively.
Wastewater source: Wastewater that was used for the study was mixed liquor that was obtained from the anaerobic zone of the same treatment plant. The mixed liquor was filtered, using Whatman No. 1 filter paper, supplemented with a particular nutrient/carbon source and then sterilized in an autoclave. To avoid the development of bacteria during the experiment, three antibiotics (penicillin, tetracycline and streptomycin) were added in concentrations of 10, 100 and 66 μg mL-1, respectively.
Nutrient/carbon supplements used: A total of nine nutrient/carbon sources were used in this study. They include: sodium acetate, sodium formate, sodium citrate, sodium succinate, peptone, tryptone, yeast extract, glucose and sucrose. They were all added singly to the mixed liquor at a concentration of 5 g L-1.
Experimental setup: To a 250 mL Erlenmeyer flask, containing 200 mL mixed liquor with a particular nutrient supplement and pH adjusted to 8.0, a rich culture of a test isolate was added and incubated in a shaker at a shaking speed of 100 rpm and incubated at 25°C.
Aliquot sample (30 mL) was removed aseptically from each flask, at the beginning and every 24 h, for the next 96 h, for the estimation of phosphate, nitrate, COD and DO, using standard method (APHA, 2001).
All reagents used were of analytical grade. Phosphate, nitrate and COD were determined using the ascorbic acid, salicylate and closed reflux methods, respectively while dissolved oxygen was determined, using a dissolved oxygen meter (model Session 8).
All experimental setups were in duplicates. An uninoculated control was always set up in all batch of experiment.
Statistical analysis: All data were analysed statistically using the SPPS computer software version 11.0. Test of significance was carried using the one-way analysis of variance (ANOVA) at 95% confidence interval.
RESULTS
Phosphate uptake in different nutrient/carbon supplements: As shown in Fig. 1, the trend in phosphate removal in the presence of the different nutrient supplements showed a significant removal in mixed liquor containing sodium acetate, glucose and sucrose. In the presence of sodium acetate, phosphate concentration decreased from 69.49 to 28.18 mg L-1, 70.14 to 24.40 mg L-1 and 68.84 to 33.26 mg L-1 in mixed liquor containing isolates A, B and C, respectively. On the other hand, mixed liquor containing glucose showed a decrease in phosphate concentration from 70.10 to 33.76 mg L-1 for isolate A, 70.83 to 27.34 mg L-1 for isolate B and 69.75 to 38.25 mg L-1 for isolate C; while a decrease from 73.26 to 42.19 mg L-1, 74.72 to 32.05 mg L-1 and 73.79 to 25.61 mg L-1 were observed in mixed liquor containing sucrose isolate A, B and C, respectively.
In general, percentage phosphate uptake of 59.45 and 65.21% were observed in the presence of sodium acetate, in isolates A and B, respectively, while percentage uptake of 65.29% was observed in the presence of sucrose for isolate C (Table 1). However, in the presence of peptone and tryptone, a phosphate release (rather than an uptake) of 3.37 and 0.60%, respectively were observed in isolate A, while in isolate B, a release of 0.33 and 4.33% were observed in the presence of sodium citrate and tryptone, respectively. In isolate C, a release of 9.69% in the presence of sodium citrate and 5.65% in the presence of peptone were observed as shown in Table 1.
Average phosphate uptake in the presence of sodium acetate, glucose or sucrose was observed to be significantly different from uptake in the presence of any of the other nutrient/carbon supplements (p≤0.05). A similar trend was observed in all the isolates.
Nitrate uptake in different carbon/nutrient supplements: The trend in nitrate uptake by the isolates revealed that in mixed liquor containing isolate A, a decrease in nitrate concentration from 11.91 to 2.13 mg L-1, 11.67 to 0.99 mg L-1, 22.50 to 6.44 mg L-1 and 30.14 to 6.74 mg L-1 was observed in mixed liquor containing sodium acetate, sodium succinate, glucose and sucrose, respectively (Fig. 2). Similarly, a decrease from 11.81 to 5.44 mg L-1 (sodium acetate), 11.82 to 1.44 mg L-1 (sodium succinate), 22.41 to 12.31 mg L-1 (glucose) and 29.74 to 10.75 mg L-1 (sucrose) were observed in mixed liquor containing isolate B, while a decrease of 12.21 to 4.15 mg L-1, 12.06 to 6.43 mg L-1, 23.46 to 4.76 mg L-1 and 29.98 to 4.54 mg L-1 were observed in the presence of sodium acetate, sodium succinate, glucose and sucrose, respectively in mixed liquor containing isolate C (Fig. 2).
Maximum nitrate uptake of 91.57 and 87.82% were observed in the presence of sodium acetate, in isolate A and B, respectively, while in the presence of isolate C, highest uptake of 84.86% was observed in the presence of sucrose. Nitrate release of 19.49, 10.75 and 22.75% were observed in the presence of tryptone, for isolates A, B and C, respectively (Table 2).
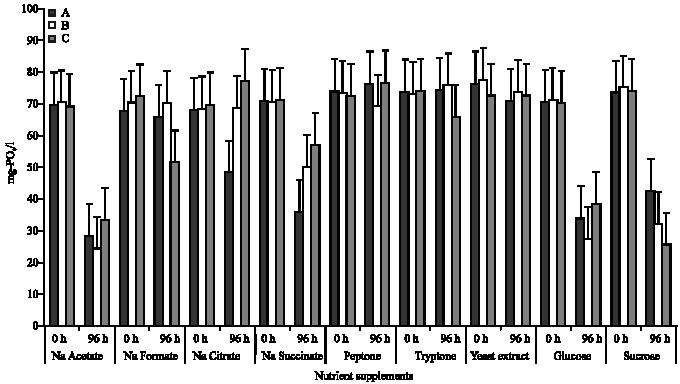 | |
| Fig. 1: | Phosphate uptake in the presence of the different nutrient supplements (A, B and C, represents the test protozoan isolates. 0 h is the beginning of the incubation while 96 h is the end of incubation) |
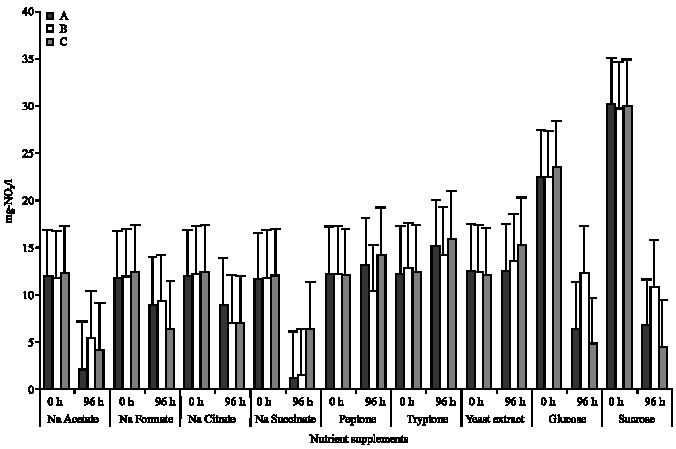 | |
| Fig. 2: | Nitrate uptake in the presence of the different nutrient supplements (A, B and C, represents the test protozoan isolates. 0 h is the beginning of the incubation while 96 h is the end of incubation) |
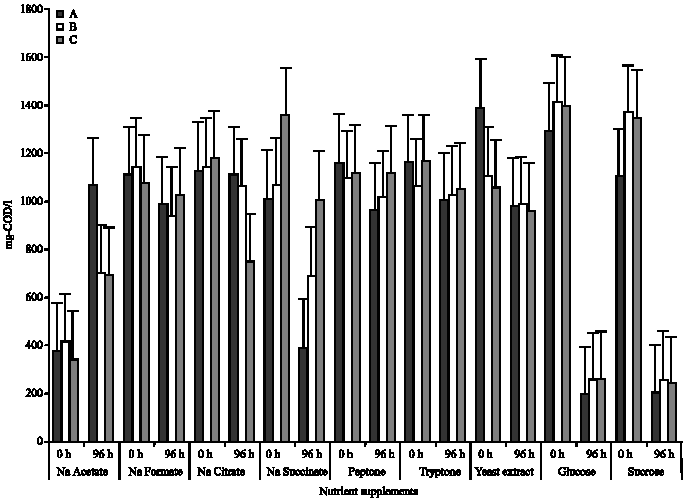 | |
| Fig. 3: | Mixed liquor COD in the presence of the different nutrient supplements A, B and C, represents the test protozoan isolates. 0 h is the beginning of the incubation while 96 h is the end of incubation) |
As was observed in phosphate uptake, average nitrate uptake by the isolates in the presence of sodium acetate, glucose or sucrose was found to be to be significantly higher than uptake in the presence of any of the other nutrient/carbon sources (p≤0.05). This observation was true for all the isolates, except isolate A were uptake in the presence of sodium succinate was found to be high also.
COD removal: In the presence of the different nutrient supplements, COD removal was remarkably observed in mixed liquor containing glucose and sucrose. This trend was irrespective of isolates. In isolate A, COD concentration of mixed liquor containing glucose and sucrose decreased from 1395.53 to 197.73 mg L-1 and 1115.95 to 201.85 mg L-1, respectively, while in isolate B and C, mixed liquor containing glucose decreased from 1407.45 to 252.18 mg L-1 and 1400.30 to 258.78 mg L-1, respectively. Also, in the presence of sucrose, mixed liquor COD decreased from 1366.20 to 508.88 mg L-1 and 1349.70 to 237.88 mg L-1 in the presence of isolates B and C, respectively (Fig. 3). This translates to a COD removal percentage of 85.83, 82.08 and 81.52% in the presence of glucose, for isolates A, B and C, respectively and 81.91, 62.68 and 82.38% in the presence of sucrose for isolates A, B and C, respectively (Table 3). In the presence of sodium acetate, a COD increase vof 64.06, 38.83 and 48.85% were observed in mixed liquor containing isolates A, B and C, respectively (Table 3).
| Table 1: | Percent phosphate uptake by the test isolates in the different nutrient supplements |
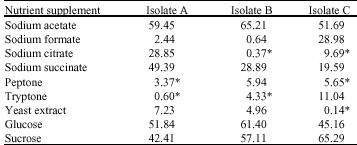 | |
| All values are averages of duplicate samples. Apart from (*) values which represent % release, all other values represent % uptake | |
| Table 2: | Percent nitrate uptake by the test isolates in the different nutrient supplements |
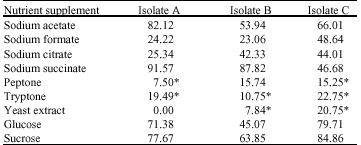 | |
| All values are averages of duplicates samples. Apart from (*) values which represent % release, all other values represent % uptake | |
| Table 3: | Percent COD removed from mixed liquor inoculated with the test isolates in the presence of the nutrient supplements |
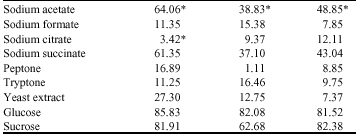 | |
| All values are averages of duplicates samples. Apart from (*) values which represent % increase, all other values represent % removal | |
| Table 4: | Concentrations of DO (mg L-1) in mixed liquor inoculated with the test isolates in the presence of the nutrient supplements |
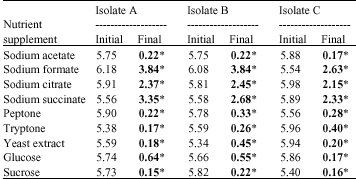 | |
| All values are averages of duplicate samples. Initial is DO concentration at the start of incubation, while final is DO concentration after 96 h incubation. (*) and bold values are the final concentrations | |
A significant COD removal in presence of either glucose or sucrose was observed to be significantly higher than removal in presence of any of the other nutrient/carbon supplements (p≤0.05). This was observation was irrespective of isolates.
DO concentration: As shown in Table 4, DO concentration decreased drastically in mixed liquor containing sodium acetate, peptone, tryptone, yeast extract, glucose and sucrose. This trend was observed in all mixed liquor containing the test isolates. However, in the presence of sodium formate, sodium citrate and sodium succinate, DO concentration was observed to have only decreased slightly. This was also similar in all the isolates.
DISCUSSION
This study revealed that the best nutrient supplement in mixed liquor for phosphate uptake by the test isolates was sodium acetate. High phosphate uptake was also observed in the presence of glucose or sucrose as nutrient supplement in mixed liquor. For nitrate uptake, sodium acetate, glucose, sucrose or sodium succinates were observed to give high uptake yield in the presence of the test isolates.
Sodium acetate has been reported to be a preferred carbon source for phosphate removal from wastewater by luxury phosphate up-taking organisms (Sang et al., 1997; Kargi and Uygur, 2003). In a study by Kargi and Uygur (2003), when investigating the effect of carbon source on biological nutrient removal in a sequencing batch reactor, their observation was, when only glucose or acetate was used as a sole carbon source, phosphate removal efficiencies were observed to be 62 and 64%, respectively. A similar trend was observed in this study. Glucose or similar carbohydrates are known to be required for energy generation to be used in poly-hydroxyl butyrate and poly-phosphate synthesis during removal (Kargi and Uygur, 2003).
In municipal wastewater treatment systems, nitrate removal is known to be normally accomplished through denitrification and normally relies on a carbon source as an electron donor (Mora et al., 2003). Gomez et al. (2000), in testing the influence of carbon source on nitrate removal of contaminated groundwater in a denitrifying submerged filter, have reported sucrose to be the least efficient carbon source in comparison with ethanol and methanol, as carbon sources. In this study, ethanol and methanol, which are likely to be toxic, were not tested as carbon sources.
Besides pH, temperature and oxygen concentration controls, carbon source dosage has been reported to be applied to submerged filter biodenitrification to contaminated water. Both simple and complex carbon sources have been employed in wastewater nutrient removal and reports have shown that addition of simple carbon sources favour nitrogen removal; hence denitrification is strongly influenced by carbon content (Barlindhaug and Odegaard, 1996; Zhe-Xue et al., 2005).
Hydrolysed molasses have been suggested as an economical and effective carbon source for nitrogen source (Zhe-Xue et al., 2005). Also, several studies have reported very good enhanced biological nutrient removal in an influent that contains significant amounts of glucose (Fukase et al., 1984; Liu et al., 1997). In a study by Guadalupe et al. (1998), they reported that in batch cultures, elimination of nitrate was enhanced when sucrose was used as a sole carbon source.
The present study revealed a high COD removal in the presence of glucose or sucrose as nutrient supplement in mixed liquor. A similar finding has been reported by Behera et al. (2007). In their report, when glucose was used as a sole carbon source during denitrification study, a near steady state was observed in percentage COD removal. This study revealed significant COD removal only in the presence of glucose, sucrose or sodium succinate as supplement in the mixed liquor. In the presence of sodium acetate, COD removal was not observed; rather there was a significant increase. COD removal efficiencies have been reported to be based on pre-treatment conditions of influent into the activated sludge system and maximum COD removal efficiency is reported to be obtained at the retention time of 15 days (Ryu et al., 2007).
Our study revealed a drastic decrease in DO concentrations of the mixed liquor in the presence of most of the nutrient supplements that we used. Earlier works have reported a decrease in DO concentrations in similar studies (Lee and Welander, 1996).
CONCLUSION
This study, which was aimed at investigating the effect of nutrient/carbon supplement on nutrient removal efficiency of three wastewater protozoan isolates, has revealed the following:
| • | That the presence of sodium acetate, sucrose or glucose as nutrient supplement in mixed liquor can enhance phosphate uptake by the test isolates. |
| • | That nitrate uptake by the test isolates is enhanced in the presence of sodium acetate, sodium succinate, glucose or sucrose as supplement in mixed liquor. |
| • | That COD removal in mixed liquor is dependent on the initial nutrient supplement in mixed liquor. |
This study will not be complete without ascertaining the optimum concentrations of the nutrient supplements that will give maximum nutrient uptake yield. This is the subject of our next investigation.
ACKNOWLEDGMENT
The authors wish to thank the National Research Foundation (NRF) of South Africa and Tshwane University of Technology for sponsoring this investigation
REFERENCES
- Akpor, O.B., M.N.B. Momba and J.O. Okonkwo, 2007. Phosphorus and nitrate removal by selected wastewater protozoa isolates. Pak. J. Biol. Sci., 10: 4008-4014.
CrossRefPubMedDirect Link - Barlindhaug, J. and H. Odegaard, 1996. Thermal hydrolysis for the production of carbon source for denitrification. Water Sci. Technol., 34: 371-378.
CrossRef - Behera, S.K., E.R. Rene and D.V. Murthy, 2007. Performance of upflow anoxic bioreactor for wastewater treatment. Int. J. Environ. Sci. Technol., 4: 247-252.
Direct Link - Caron, D.A., C.G. Joel and M.R. Dennette, 1998. Experimental demonstration of the roles of bacteria and bacterivorous protozoa in plankton nutrient cycles. Hydrobiologia, 159: 27-40.
CrossRef - Coleman, D.C., 1994. The microbial loop concept as used in terrestrial soil ecology studies Microbial. Ecol., 28: 245-250.
CrossRef - Foy, R.H. and P.J.A. Withers, 1995. The contribution of agricultural phosphorus to eutrophication. Proceedings of the Fertilizer Society, (FS'95), Belfast, UK., pp: 1-32.
Direct Link - Fukase, T., M. Shibata and Y. Miyaji, 1984. The role of an anaerobic stage on biological phosphorus removal. Water Sci. Technol., 17: 69-80.
Direct Link - Gomez, M.A., J. Gonzalez-Lopez and E. Hontoria-garcia, 2000. Influence of carbon source on nitrate removal of contaminated groundwater in a denitrifying submerged filter. J. Hazard. Mater., 80: 69-80.
Direct Link - Pinar, G., K. Kovarova, T. Egli and J.L. Ramos, 1998. Influence of carbon source on nitrate removal by nitrate-tolerant Klebsiella oxytoca CECT 4460 in batch and chemostat cultures. Applied Environ. Microbiol., 64: 2970-2976.
Direct Link - Kargi, F. and A. Uygur, 2003. Effect of carbon source on biological nutrient removal in a sequencing batch reactor. Bioresour. Technol., 89: 89-93.
Direct Link - Kargi, F., A. Uygur and H.B. Sava, 2005. Phosphate uptake and release rates with different carbon sources in biological nutrient removal using a SBR. J. Environ. Manage., 76: 71-75.
Direct Link - Kross, B.C., G.R. Hallberg, D.R. Bruner, K. Cherryholmes and J.K. Johnson, 1993. The nitrate contamination of private well water in Iowa. Am. J. Public Health, 83: 270-272.
CrossRefDirect Link - Lee, N.M. and T. Welander, 1996. Use of protozoa for decreasing sludge production in aerobic wastewater treatment. Biotechnol. Lett., 18: 429-434.
Direct Link - Liu, W., K. Nakamura, T. Matsuo and T. Mino, 1997. Internal energy-based competition between polyphosphate-and-glycogen-accumulating bacteria in biological removal reactors: Effect of P/C feeding ratio. Water Res., 31: 1430-1438.
CrossRef - Mora, F.R., Ferrara, G. de Giner, A.R. Andara and J.L. Esteban, 2003. Effect of organic carbon shock loading on endogenous denitrification in sequencing batch reactors. Bioresour. Technol., 88: 215-219.
Direct Link - Ryu, H., S. Lee and K. Chung, 2007. Chemical oxygen demand removal efficiency of biological treatment process treating tannery wastewater following seawater flocculation. Environ. Eng., 24: 394-399.
Direct Link - Sang, L., P. Jang-Ho, K. Kwang-Baik and K. Ben, 1997. Effect of fermented swine wastes on biological nutrient removal in sequencing batch reactors. Water Res., 31: 1807-1812.
CrossRef - Zhe-Xue, Q., S.J. Yin, R.Y. Chang, J.L. Jay and T.L. Sung, 2005. Hydrolysed molasses as an external carbon source in biological nitrogen removal. Bioresour. Technol., 96: 1690-1695.
Direct Link