
Abdelhamid M. Hamad
Institute of Biological Science, Faculty of Science, University of Malaya, 50603, Kuala Lumpur, Malaysia
R.M. Taha
Institute of Biological Science, Faculty of Science, University of Malaya, 50603, Kuala Lumpur, Malaysia
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 22 | Page No.: 4180-4185







ABSTRACT
The effect of 6-benzylaminopurine (BAP) at concentration range of 0 to 3.75 mg L-1 applied at concentration difference of a fixed increment of 0.25 mg L-1 on the in vitro shoot formation and average and total of shoot length and weight per explant of smooth cayenne pineapple were evaluated. BAP significantly affected the average of shoot number, length and weight but did not affect the total length and total weight per explant. The maximum shoot formation (12 shoots/explant) occurred at 4 concentrations of BAP (1.75, 2.0, 2.25 and 3.5 mg L-1) while tallest (25 mm) and heaviest (0.60 g) shoots obtained on MS hormone free medium. Shoot formation pattern showed two sudden increases in shoot formation. The first occurred at 1.25 and second at 3.50 mg L-1 and two sharp declines at 2.5 and 3.75 mg L-1. Identifying of the concentrations with contrasting effect where an increase and decline in the shoot formation pattern occur provide an essential base from which an informative successful histological and biochemical studies could be designed to elucidate shoot formation mechanism.
PDF Abstract XML References Citation
How to cite this article
Abdelhamid M. Hamad and R.M. Taha, 2008. Effect of Benzylaminopurine (BAP) on in vitro Proliferation and Growth of Pineapple (Ananas comosus L. Merr.) cv. Smooth Cayenne. Journal of Applied Sciences, 8: 4180-4185.
DOI: 10.3923/jas.2008.4180.4185
URL: https://scialert.net/abstract/?doi=jas.2008.4180.4185
DOI: 10.3923/jas.2008.4180.4185
URL: https://scialert.net/abstract/?doi=jas.2008.4180.4185
INTRODUCTION
In vitro multiplication of pineapple shoots could be achieved in medium enriched with BAP alone (Be and Debergh, 2006), combination of two hormones, BAP plus, naphthalene acetic acid (NAA) (Firoozabady and Gutterson, 2003), indole acetic acid (IAA) (Hamad and Taha, 2008), indole butyric acid (IBA) (Boxus et al., 1991) and 2,4-dichlorophenoxy acetic acid (2,4-D) (Liu et al., 1989) and combination of three hormones, BAP plus two auxins as NAA and IBA (Soneji et al., 2002), NAA and IAA (Mathews and Rangan, 1979) and IAA and IBA (Teixeira et al., 2006). Single application of BAP is simpler and cheaper and should be favored over combination of two and three hormones. However, the optimum concentration of BAP was not agreed upon. BAP at 1.0 (Be and Debergh, 2006), 1.5 (Almeida et al., 2002), 2.0 (Bhatia and Ashwath, 2002), 2.5 (Smith et al., 2002), 3.0 (Firoozabady and Gutterson, 2003) and 4.0 mg L-1 (Omokoio et al., 2001) were recommended for multiplication of pineapple. It seemed that the optimal BAP concentration lay between 1.0 and 4.0 mg L-1. However, in all of these studies either the tested concentrations were as low as 2 concentrations (Almeida et al., 2002) or the difference among the tested concentrations range (the increment) was too large, 2.0 mg L-1 (Omokoio et al., 2001) or both to draw a conclusive conclusion. While BAP concentration up to 1.0 mg L-1 was thoroughly investigated at a concentration differences of 0.2 (Be and Debergh, 2006) and 0.25 mg L-1, (Aydieh et al., 2000) a concentration difference of 0.5 mg L-1 used for testing concentrations range up to 1.5 (Zepeda and Sagawa, 1981), 2.0 (Fernando, 1986), 2.5 mg L-1 (Bhatia and Ashwath, 2002). Concentration up to 3 mg L-1 tested at concentration difference of 1.5 mg L-1 (Almeida et al., 2002) and up to 12 mg L-1 at concentration difference of 2.0 mg L-1 (Omokoio et al., 2001).
Being about 30 different hormone treatments have been reported for multiplication of pineapple, raise the question of which should be used. Interestingly, although in most of these treatments, whether the hormones applied singly or in combinations, the difference was concentration more than the types of hormones, the effect of BAP concentrations above 1.0 mg L-1 on pineapple cultures were roughly investigated (few levels and large increment). It is well known that the same hormone could promote at one concentration and inhibit at other and a minute difference could draw the line between the two effects. Hence, it is highly likely that the best concentration was left out and not being included in these studies of pineapple. In fact this case was proven to be true for caster bean. Using wider concentration range enabled Sujatha and Reddy (1998) to select treatment that increased castor proliferation rate five times higher than previously reported. In addition, due to using of few concentrations and either unequal or too large increments, the shoot formation pattern in response to different BAP in all of the previous studies of pineapple was neither discussed nor it could be visualized from the reported data. We think that this area needs to be covered not only for optimization of multiplication but most importantly for pin point of the concentrations which would have contradicting effect on the shoot formation pattern. From there further studies could be conducted to elucidate the mode of hormone effect and mechanism of shoot formation.
The objectives of this study were first to test the effect of singly applied BAP at a wider range of 0 to 3.75 mg L-1 and smaller increment of 0.25 mg L-1 than those reported before on the shoot formation per explant. Secondly, to evaluate the amount of growth in term of average length and weight of the produced shoots and total length and weight that could be obtained from single explant for assessment of biomass production. Thirdly, to follow the shoot formation pattern and determine the concentrations at which the BAP promote and inhibit the proliferation and growth of pineapple and how much change would occur in response to one or several increase of a fixed increment of 0.25 mg L-1 of BAP.
MATERIALS AND METHODS
Three liters of MS medium (Murashige and Skoog, 1962) were prepared from stock solutions. The medium divided into 16 beakers (180 mL each). No hormone added to the first beaker and BAP at 0.25, 0.50, 0.75, 1.00, 1.25, 1.50, 1.75, 2.00, 2.25, 2.50, 2.75, 3.00, 3.25, 3.50 and 3.75 mg L-1 were added to beakers No. 2-16, respectively. The medium pH adjusted to 5.7 and the content of each beaker dispensed equally (20 mL) into 9 glass jars (5x15 cm). Agar at 7 g L-1 was added to each jar, the jar closed with autoclavable lid and the medium autoclaved at 121 °C and kg cm-2 for 25 min. Stock cultures that were maintained by subculturing every 75 days on agar solidified MS medium enriched with BAP at 2.3 mg L-1 were used as explants source for this study. One shoot was cultured per each jar under aseptic conditions. The cultures incubated under a constant temperature of 25 °C ± 2 and 16 h of artificial light provided by fluorescent lamp. After 60 days, the cultures were taken of the incubation room and the multiple shoots were picked up, weighted and separated into individual shoots for counting number and measuring the shoot length. The weight of multiple shoots per each culture considered as total weight per explant. The length of shoots summed and considered as total length per explant. Average weight and length of shoots computed from dividing total weight and total length by number of shoots per explant. Each treatment consisted of nine cultures (3 cultures per replicate) and the data recorded from each three of the nine cultures summed and divided by 3 to obtain the average value for each replicate. Analysis of variance, treatments means separation by Duncan Multiple Range Test, correlation and regression analysis was done at p<0.5 using SPSS 11.0 statistical package.
RESULTS
Table 1 shows that the shoots formation increased in response to every 0.25 mg L-1 increase in concentration of BAP up to 1.0 mg L-1 at almost steady rate (1 shoot/each 0.25 mg L-1 increase in concentration). Then an increase of 0.25 mg L-1 resulted in a sudden surge in shoot formation and the shoot number rose from 6 to 10 shoots/explant as the concentration rose from at 1.0 to 1.25 mg L-1. After that, the rate of shoot formation remained almost stable around 11 shoots per explant for four consecutive increases in the concentration of BAP by 0.25 mg L-1 till the concentration reached 2.25 mg L-1. Then an increase of 0.25 mg L-1 caused a sharp decline in shoot formation and the shoots number decreased from 12 to 8 shoots/explant in response to increase of BAP concentration from 2.25 to 2.5 mg L-1.
| Table 1: | Effect of BAP concentrations on shoots No., average shoot length (Lx), shoot weight (Wx) and total shoots length (LT) shoots weight (WT) per explant of smooth cayenne pineapple |
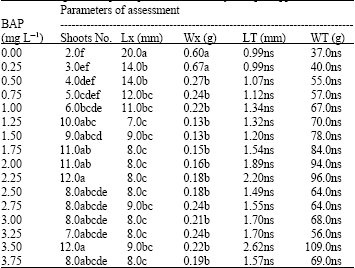 | |
Number represent means of 9 shoot buds individually cultured in 20 mL of agar solidified (7 g L-1) MS medium containing 30 g L-1 of sucrose and enriched with BAP at different concentration (mg L-1) for 60 days. pH adjusted to 5.7. Means of the same column followed by same letter(s) were not significantly different at p = 0.05 using Duncan`s multiple range test. ns (No significant effect on total weight (p = 0.2496) and total length (p = 0.2135) | |
| Table 2: | Correlation coefficient and regression equations among BAP at concentration range of 0.0-3.75 mg L-1 and the growth parameters of smooth cayenne pineapple cultured on agar solidified full strength MS medium for 60 days |
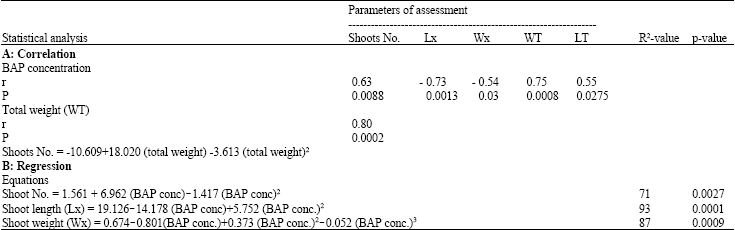 | |
Lx: Average length of shoot, LT: Total length of shoots per explant, Wx: Average fresh weight of shoot, WT: Total fresh weight of shoots per explant | |
The rate again remained stable at 8 shoots per explant for four consecutive increases in BAP concentration by 0.25 mg L-1 till the concentration reached 3.25 mg L-1. After that increase of concentration by 0.25 mg L-1 from 3.25 to 3.5 mg L-1 reversed the shoot pattern and a second surge of shoot formation from 7 to 12 shoots per explant occurred. However, the potential for shoot formation the explants had just gained was lost upon raising the concentration to 3.75 mg L-1. Generally, there were two sharp increases and decreases in shoot formation. The first increase occurred as the concentration raised from 1.0 to 1.25 mg L-1 BAP and the second increase as the concentration raised from 3.25 to 3.5 mg L-1 while the first decrease occurred as the concentration raised from 2.25 to 2.5 and the second decrease when the concentration raised from 3.5 to 3.75 mg L-1. There were no significant difference between shoot number per explant at 1.75, 2.0, 2.25 and 3.5 mg L-1 of BAP and all induced the highest shoot formation (12 shoots/explant) and no significant differences between shoot number per explant at 2.5, 2.75, 3.0 and 3.75 mg L-1 of BAP and all resulted in intermediate shoot formation (8 shoots per explant). At any concentration below 1.0 mg L-1, the shoot formation was less than 6 shoots per explants and each increase of 0.25 mg L-1 resulted in significant increase in number of shoots per explant. Regression analysis (R2 = 0.71) indicated that 71% of the difference in shoots formation was caused by differences in the concentration of BAP and the relation between BAP concentration and shoot formation was cubic polynomial relationship (Table 2). Correlation analysis indicated that the relation between shoot formation and BAP concentrations was positive (r = 0.63) and the shoot increased as the BAP concentration increased but was negative with average length (r = -0.83) and weight (r = -0.76).
The tallest (25 mm) and heaviest (0.67 g) shoots obtained in hormone free and at 0.25 mg L-1 and the shortest (7 mm) and lightest (0.13 g) shoots at 1.25 mg L-1. The shoot length decreased as the BAP concentration increased up to 0.5 mg L-1 while shoot weight decreased as concentration increased up to 1.25 mg L-1. Concentrations above 0.5 and 1.25 up to mg L-1 did not cause any significant difference in average shoot length and shoot weight/explant respectively (Table 1). Regression coefficient of shoot length (R2 = 0.93) and shoot weight (R2 = 0.87) indicated that 93% of differences in shoot average length and 87% of the differences on average shoot weight were due to BAP concentrations (Table 2). Correlation analysis between BAP concentrations and both of average shoot length (r = -0.73) and average shoot weight (r = -0.54) indicated a negative relationship. Both of the shoot length and weight average correlated positively with each other (r = 0.82) and negatively (r = -83 and -76, respectively) with shoot formation. The effect of different BAP concentrations on average shoot length and average weight per explant seemed to follow a similar pattern. Both responded negatively and only to low concentration range (0.25 to 1.25 mg L-1) of BAP.
BAP concentrations, on the other hand, had no significant effect on total length (p<0.2135) and total weight (p<0.2496) of shoots per explant. Nevertheless, the largest total length (109 mm) and heaviest total weight (2.61 g) obtained on medium enriched with BAP at 3.5 mg L-1 and the lowest total length (37 mm) and total weight (0.99 g) obtained on hormone free medium (Table 1). The total length and total weight of shoots showed similar pattern but opposite to the pattern of the average shoot length and weight per explant. Both of the total length and weight were not affected by BAP concentrations, correlated positively with shoot formation (r = 93 and 80) and with each other (r = 85) and negatively with average shoot length (r = -66 and -58) and with average shoot weight (r = -66 and 45).
The optimal concentration of BAP for multiplication of pineapple (Ananas comosus L. Merr.) cv. smooth cayenne in solid MS appeared to be 1.75, 2.0, 2.25 and 3.5 mg L-1. Each resulted in the highest number of shoot formation (12 shoots/explant) and equal shoot length of 8 mm long and about 0.18 g/explant (average weight/explant) and over 1.5 g per culture (total weight/explant) per 60 days of incubation.
DISCUSSION
The main goal of tissue culturist is to optimize the multiplication. However, in most of the time they follow try and error approach to select best concentration of a one hormone or best hormone type applied singly or in combinations. Sujatha and Reddy (1998) used this approach to optimize the multiplication of castor bean and come up with hormone treatment that resulted in multiplication five times higher than previously reported treatments. The 12 shoots per explant of pineapple obtained in this study in agar solidified MS enriched with BAP at 1.75, 2.25 and 3.5 mg L-1 is higher than the previously reported 10 (Sripaoraya et al., 2003) and 7 shoots per explant (Bhatia and Ashwath, 2002) obtained in response to BAP at 2.0 mg L-1 and the 9 (Be and Debergh, 2006), 7 (Aydieh et al., 2000) and 3 shoots per explant (Zepeda and Sagawa, 1981) obtained in response to BAP at 1.0 mg L-1 and the 10 shoots (Firoozabady and Gutterson, 2003) in response to 3.0 mg L-1. The result did not support the use of BAP at 1.5 mg L-1 suggested by Ameida et al. (2002) and 2.5 mg L-1 suggested by Smith et al. (2002).
Although trial and error approach is very important and the most common used approach, the most optimal treatment would not be achieved unless the mode of hormone action and shoot formation mechanism is elucidated. Understanding of mode of action and mechanism require histological and chemical analysis of the endogenous content of the explants in response to different concentrations and type of hormone. The success on this approach depends mainly on proper selection of the concentration of the added hormone and time at which these analyses should be taken. Hence, the first essential step is to determine the growth and shoot formation pattern. Surprisingly in most of pineapple studies, shoot formation pattern in response to a continuous increase in concentrations by one fixed unit of concentration over wide range were either totally ignored or it could not be discern from the limited reported data. Exogenously applied hormone stimulates synthesis of endogenous hormones in pineapple leaf culture and histological and chemical studies confirmed strong association between sharp increase in the endogenous IAA (indole acetic acid) and 2iP (isopentenyladenine) content and formation of shoots (Hamasaki et al., 2005; Mecier et al., 2003). In these studies, a fixed and arbitrary selected hormone concentration was used. If two different concentrations one inhibit and other promote shoot formation were used the results would be more informative. In this study we think that the first step has been taken, the changes in shoot formation were followed at fixed unit of concentration increase (0.25 mg L-1) and the different concentrations at which the BAP promote and inhibit the shoot formation have been identified.
It is very interesting to note that a 0.25 mg L-1 difference in concentration of BAP could reverse the mode on either positive or negative direction and the effect seemed to follow symmetrical trend. Raising the concentration by 0.25 mg L-1 from 1.0 to 1.25 and from 3.25 to 3.5 resulted in sudden sharp increase in the shoot formation capacity by 4 more shoots (30% more) and the capacity just gained maintained for 4 consecutive 0.25 mg L-1 increases in concentration to be lost by the fifth. Raising the concentration by the same amount (0.25 mg L-1) from 2.25 to 2.5 and from 3.5 to 3.75 mg L-1 caused on the contrary drastic loss of 4 shoots and the loss on the capacity persisted for 4 consecutive increases in the concentration to be reversed by the fifth. It appeared that 4 consecutive increases in concentration by 0.25 mg L-1 are required before the fifth could cause sudden sharp increase or drastic decrease in shoot formation. Nielsen et al. (1995) proposed one receptor and two binding sites model for cytokinin action in plant cell. Functioning receptor-hormone complex is the one in which the sites occupied by the endogenous hormones. High concentrations of exogenous hormone compete with endogenous hormone for the active sites or change the conformation of the receptor and reduce the total number of functioning complex. The ratio between the exogenous and endogenous hormone and the natural turn over of functional complex determine the shoot formation pattern. The shoot formation pattern could also relate to changes in the other chemical content within the explants and the different concentrations of the hormone may affect the uptake of nutrient and medium pH differently. Chemical analysis of agar-solidified medium was successfully used for identification of anti-hyperhydricity factors and facilitates commercial micropropagation of radiata pine (Nairn et al., 1995). Similar approach could be proven successful for pineapple. Comparing of histological and chemical analysis of explants and MS components in medium enriched with BAP at 1.25 and 3.5 with that from MS enriched with BAP at 2.5 and 3.75 would probably shed some light on the physiology of in vitro shoot formation and growth of pineapple.
The pineapple tissue culture is usually done for propagation purpose and the best treatment judged by the rate of shoot formation. Other parameters such as total or average fresh weight are rarely reported. As the goal was propagules production, neglecting of weight are understandable. However, since there is possibility for extraction of bromelain for medical purpose and use of biomass for animal feeding reporting of weight would be more important than any other parameters. The heaviest shoot (average weight per shoot) obtained in medium contained BAP at 0.25 mg L-1 but there were no significant different in the total weight per explant (weight/culture). Hence, for biomass production hormone addition is not required. On the contrary, Pereze et al. (2003) obtained higher fresh weight, protein content and protease activity on response to BAP at 0.5 mg L-1. Be and Debergh (2006) reported that although BAP at 1.0 mg L-1 and combination of BAP at 1.0 plus IBA at 0.5 mg L-1 induced equal number of shoots, the second treatment doubled the total fresh weight/culture. Existing of positive correlation between shoot numbers, total weight, total length (Table 2) indicated that a system for optimum production of both propagules and biomass could be developed using MS medium enriched with proper concentration of BAP. In addition, in cases when high explants density are used and in large scale production counting of individual shoot is not practical. The high correlation between number and total weight of shoots (r = 0.80) indicated that rather than individual counting, the shoot number could possibly estimation as function of total weight according to the model presented (Table 2). Firoozabady and Gutterson (2003) counted the number of shoots in specific unit of weight and used that ratio for estimation of shoot number from a total weight obtained in bioreactor system. Salehi and Khosh-Khui (1997) suggested a model by which the shoot length after one week in culture could be used for estimation of the number of shoots that miniature rose would produce after four weeks of incubation.
REFERENCES
- De Almeida, W.A.B., G.S. Santana, A.P.M. Rodriguez and M.A.P. Carvalho Costa, 2002. Optimization of a protocol for the micropropagation of pineapple. Rev. Bras. Frutic., 24: 296-300.
CrossRef - Be, L.V. and P.C. Debergh, 2006. Potential low-cost micropropagation of pineapple (Ananas comosus). S. Afr. J. Bot., 72: 191-194.
CrossRef - Bhatia, P. and N. Ashwath, 2002. Development of rapid method for micropropagation of a new pineapple (Ananas comosus (L.) Merr. clone Yeppoon gold. Acta Hortic., 575: 125-131.
Direct Link - Boxus, P., J.M. Terzi, C. Lieves, M. Pylyser, P. Ngaboyamahina and K. Duhem, 1991. Improvement and perspectives of micropropagation techniques applied to some hot climate plants. Acta Hortic., 289: 55-59.
Direct Link - Firoozabady, E. and N. Gutterson, 2003. Cost-effective in vitro propagation methods for pineapple. Plant Cell Rept., 21: 844-850.
Direct Link - Hamad, A.M. and R.M. Taha, 2008. The effect of different hormones and incubation periods on in vitro proliferation of pineapple (Ananas comosus L.) Merr cv. Smooth Cayenne) shoot-tip culture. Pak. J. Biol. Sci., 11: 386-391.
CrossRefPubMedDirect Link - Hamasaki, R.M., E. Purgatto and H. Mercier, 2005. Glutamine enhances competence for organogenesis in pineapple leaves cultivated in vitro. Braz. J. Plant. Physiol., 17: 383-389.
CrossRefDirect Link - Mathews, V.H. and T.S. Rangan, 1979. Multiple plantlets in lateral bud and leaf explant in vitro culture of pineapple. Sci. Hortic., 11: 319-328.
CrossRef - Mercier, H., B.M. Souza, J.E. Kraus, R.M. Hamasaki and B. Sotta, 2003. Endogenous auxin and cytokinin contents associated with shoot formation in leaves of pineapple cultured in vitro. Braz. J. Plant. Physiol., 15: 107-112.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nairn, B.J., R.H. Furneaux and T.T. Stevenson, 1995. Identification of an agar constituent responsible for hydric control in micropropagation of radiata pine. Plant Cell Tiss. Org. Cult., 43: 1-11.
CrossRef - Omokoio, N.D., M.A. Tita and N. Niemenak, 2001. Direct in vitro regeneration of Ananas comosus (L.) Merrill var. Cayenne from crowns grown in liquid medium. Fruits, 56: 415-421.
CrossRefDirect Link - Perez, A., L. Napoles, J.C. Lorenzo and M. Hernandez, 2003. Protease excretion during pineapple micropropagation in temporary immersion bioreactors. In vitro Cell Dev. Biol.-Plant, 39: 311-315.
CrossRef - Smith, M.K., H.L. Ko, S.D. Hamill and G.M. Sanewski, 2002. Pineapple transformation: Mangging somaclonal variation. Acta Hortic., 575: 69-74.
Direct Link - Soneji, J.R., P.S. Rao and M. Mhatre, 2002. Somaclonal variation in micropropagated dormant axillary buds of pineapple (Ananas comosus L. Merr.). J. Hort. Sci. Biotechnol., 77: 28-32.
Direct Link - Sripaoraya, S., R. Marchant, J.B. Power and M.R. Davey, 2003. Plant regeneration by somatic embryogenesis and organogenesis in commercial pineapple (Ananas comosus L.). In vitro Cell Dev. Biol.-Plant, 39: 450-454.
CrossRef - Sujatha, M. and T.P. Reddy, 1998. Differental cytokinin effects on the stimulation of in vitro shoot proliferation from meristematic explants of castor (Ricinus communis L.). Plant Cell Rept., 17: 561-566.
CrossRef - Teixeira, S.L., J.M. Ribeiro and M.T. Teixera, 2006. Influence of NaClO on nutrient medium sterilization and on pineapple (Ananas comosus cv Smooth cayenne) behavior. Plant Cell Tissue Org. Cult., 86: 375-378.
CrossRefDirect Link - Nielsen, J.M., J. Hansen and K. Brandt, 1995. Synergism of thidiazuron and benzyladenine in axillary shoot formation depends on sequence of application in Miscanthus X ogiformits Gigantenus. Plant Cell Tissue Org. Cult., 41: 165-170.
CrossRef