
Theodosy Msogoya
Postgraduate School, Writtle College, Chelmsford CM1 3RR, UK
Brian Grout
Life Sciences, University of Copenhagen, 2630-Taastrup, Denmark
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 15 | Page No.: 2703-2710







ABSTRACT
This study was conducted to investigate black sigatoka disease and drought stress responses of off-type plants derived from shoot-tip micropropagation of East African highland banana (Musa AAA East Africa) landrace Uganda. Results showed that the off-type plants were more (p<0.05) tolerant to black sigatoka disease with the infection index of 17.5% compared to 30.1 and 22.8% of the micropropagation (MP) derived phenotypically normal plants and Conventional Propagation (CP) derived plants with no tissue culture history in their ancestry, respectively. On the contrary, the off-type plants were more (p<0.05) vulnerable to water stress with leaf senescence of 87.7% at soil water deficit of 630 millibars. The leaf senescence of the MP and CP derived plants at the same soil moisture deficit was 79.5 and 66.7%, respectively. During this stress period each off-type plant produced one sucker, while the true-to-type plants were unable to do so. Leaf structural analysis revealed that the off-type plants had higher (p<0.05) stomatal density of 16.0 mm-2 of the upper leaf surface. Conversely, the MP and CP derived plants had each 12.3 and 11.0 stomata mm-2 of the leaf upper surface. Similarly, the off-type plant leaves were more hydrophobic with higher (p<0.05) epicuticular waxiness of 684.6 μg cm-2. The epicuticular wax content of the MP and CP derived plant leaves was as low as 646.2 and 647.7 μg cm-2, respectively. The water stressed off-type plant leaves exhibited higher (p<0.05) membrane damage with ion leakage of 168.2 μS cm-1 compared to 139.7 and 136.8 μS cm-1 of the MP and CP derived plants. Moreover, the water stressed leaves of the off-type plants had enhanced total antioxidant activity of 5.17 M trolox equivalent per milligram proteins, whereas the total antioxidant activity of the MP and CP derived plant leaves was as low as 3.76 and 3.67 M trolox equivalent per milligram proteins, respectively.
PDF Abstract XML References Citation
How to cite this article
Theodosy Msogoya and Brian Grout, 2008. Altered Response to Biotic and Abiotic Stress in Tissue Culture-Induced Off-Type Plants of East African Highland Banana (Musa AAA East Africa). Journal of Applied Sciences, 8: 2703-2710.
DOI: 10.3923/jas.2008.2703.2710
URL: https://scialert.net/abstract/?doi=jas.2008.2703.2710
DOI: 10.3923/jas.2008.2703.2710
URL: https://scialert.net/abstract/?doi=jas.2008.2703.2710
INTRODUCTION
A controlled plant response to tissue culture-mediated stress may result in an enhanced plant protection against adverse biotic and abiotic environment (Cassells and Curry, 2001). Good examples include, in vitro derived off-type banana plants with tolerance to yellow sigatoka (Mycosphaerella musicola Lench) (Trujillo and Garcia, 1996) and somaclonal variants from African plantains with tolerance to black sigatoka (Mycosphaerella fijiensis Morelet) (Nwauzoma et al., 2002). Also reported are somaclonal variants of wheat with tolerance to high aluminium level (Carver and Johnson, 1989) and Cavendish banana cv. Grand nain with tolerance to mild winter (Damasco et al., 1996). On the contrary, a high incidence of banana streak virus was reported in micropropagated plantain cv. Superplatano (Krikorian et al., 1999) and severe leaf spot diseases (Ascochyta rhei and Ramularia rhei) in micropropagated rhubarb line CP49 (Zhao, 2004).
Tissue culture-mediated stress enhances the production of free radicals, which attack unsaturated lipids in the cell membrane and other macroelements such as nucleic acids and proteins (Benson, 2000; Campos et al., 2003; Johnston et al., 2006). In response to this stress, plants produce more antioxidants to defend themselves against the toxicity of reactive oxygen species and their secondary products (Prochazkova et al., 2001). In addition, tissue culture conditions often affect the formation of leaf waxiness, number of stomata and guard cell functioning (Blanke and Belcher, 1989; Jonathan et al., 1997; Majada et al., 1998). Changes in cell membrane stability, antioxidant activity, leaf waxiness and stomatal density affect plant responses to ex vitro biotic and abiotic stresses (Marin et al., 1988; Kirdmanee et al., 1996; Busogoro et al., 2004).
Shoot-tip micropropagated East African highland banana (Musa AAA East Africa) landrace Uganda exhibited high incidence of off-type plants. This instability could be utilised as a tool for the improvement of landrace Uganda if the agronomic performance of the off-type plants were known. The agronomic improvement of landrace Uganda, like all landraces of East African highland bananas, is required to increase plant tolerance to various biotic and abiotic stresses (Rukazambuga et al., 1998). The objective of this study was to investigate the tolerance to drought stress and black sigatoka disease of tissue culture-derived off-type plants of landrace Uganda.
MATERIALS AND METHODS
Description of study areas and plant materials: East African highland banana landrace Uganda with no micropropagation history in their ancestry was micropropagated at Sokoine University of Agriculture according to Maerere et al. (2003). Briefly, initial explants of 2.0X2.5X50 mm were isolated from sword suckers, surface-sterilised in ethanol (96% v/v) for 2 min and disinfected for 20 min in 3.5% (m/v) sodium hypochloride. The cubes were rinsed three times in sterile distilled water and aseptically dissected to remove bleached tissues. A final shoot tip explant of about 10X10X15 mm consisted of a portion of the corm, 3-4 leaf primordia and meristematic dome. The growth media comprised Murashige and Skoog (1962) salts and vitamins at 4.4 g, L-cystein at 40.0 mg, 6-benzylaminopurine at 6.0 mg, indole-3-acetic acid at 2.0 mg, sucrose at 30.0 g and agar at 8.0 g L-1. The cultures were initially incubated in darkness at 26-30°C for 4 weeks. The proliferation was carried out using the same growth media and temperatures and a photoperiod of 16 h day-1 supplied by cool white fluorescent tubes. Successive subcultures were carried out at an interval of 4 weeks.
The shoots were transferred to a rooting media at the fifth subculture. The rooting media consisted of MS salts and vitamins at 4.4 g, L-cystein at 40.0 mg, naphthaleneacetic acid at 2.0 mg, sucrose at 30.0 g and agar at 8.0 g L-1. Rooted plantlets at about 5 cm tall were acclimatised in coconut peat media for 4 weeks under shades of 50 and 30%, respectively. The in vitro suckers were planted in the field at Sokoine University of Agriculture in May 2004. Planting holes of 100X100X100 cm dimension spaced at 300 m apart were prepared and filled each with 60 L of farmyard manure. The crop received optimal management, including weeding, irrigation during dry season, application of farmyard manure in subsequent years and desuckering to maintain four plants per stool. Off-type plants were detected and 30 suckers each of the off-type banana (treatment), micropropagation (MP) derived phenotypically normal banana (control 1) and Conventional Propagation (CP) derived banana with no tissue culture history in its ancestry (control 2) were collected and planted in new plots in May 2006.
Black sigatoka disease: Black sigatoka disease infection under natural inoculation conditions was assessed using 30 field-grown plants of the off-type, MP and CP derived plants in April 2007. The disease was evaluated during the rainy season using fully expanded leaves based on 0-6 severity scale according to Gauhl et al. (1993). In this scale, 6 was scored when 51-100% of the total leaf area died as a consequence of the disease infection and 0 when no disease symptom was visually observed on the leaf. The disease infection index was calculated according to Orjeda (1998).
Drought stress responses: Suckers of the off-type, MP and CP derived plants were collected from the field-grown parents and planted in 20 L containers in February 2007. The growth media consisted of forest soil, farmyard manure and rice husk at 3:2:1 (v/v), respectively. Thirty container plants at 1.5 m tall stage each of the off-type, MP and CP derived banana were transferred to a plastic tunnel with 30% shade screen and average day temperatures, relative humidity and light of 26°C, 68% and 2,000 lux, respectively. In April 2007, the plants were water-stressed by withholding irrigation until the soil moisture level monitored by a tensiometer (Eijkelkamp Agrisearch, The Netherlands) reached 630 millibars. The plant water stress response was evaluated based on the decrease in pseudostem diameter, leaf senescence and sucker production. The pseudostem diameter at 65 cm high was measured using a vernier caliper, whereas leaf senescence was scored when at least a quarter of one or both side(s) of the margin length was yellow. Suckers were counted as soon as they emerged out to the soil surface.
Stomatal density: The leaf stomatal density was determined from banana cigar leaves according to Marin et al. (1988). Stomatal density was calculated as the number of stomata per unit leaf area of the leaf lower and upper surfaces.
Leaf epicuticular waxiness: The wettability of leaf upper and lower surfaces was estimated based on static contact angle according to Koch et al. (2006). The contact angle between water droplet and leaf surface was measured using a protractor. Similarly, leaf waxiness was determined gravimetrically according to Kim et al. (2006). Leaf waxiness was expressed in microgram per leaf area (total of upper and lower surfaces).
Electrolyte leakage: Electrolyte leakage was determined using fully expanded leaves according to Saneoka et al. (2004). A pilot study was carried out to determine mannitol concentration that resulted in a highest leaf electrolyte leakage. Ten millilitres of mannitol solution at 0.1, 0.2, 0.3, 0.4 and 0.5 M was each dispensed in a test tube. The initial conductivity of the mannitol solutions in the test tubes was measured at 25°C using a conductimeter (Hanna Instruments Limited). Six leaf discs from washed leaves were dipped in the mannitol solution in each test tube and incubated at 26°C for 3.0 h on a rotary shaker at 150 rpm. The final conductivity of the solution was measured again and the electrolyte leakage was calculated as the difference between the final and initial conductivity of the mannitol solution. The highest ion leakage was observed at 0.3 M of mannitol. Leaf discs of equal surface area from the off-type, MP and CP derived plants were dehydrated at 0.3 M mannitol solution as described above. Electrolyte leakage was measured as described above and the ion leakage was expressed in microsiemen per centimetre.
Antioxidant activity: Cigar leaves from the off-type, MP and CP derived plants were collected in the morning, wrapped in moist tissue papers and air-transported to the United Kingdom. The leaves were kept for 5 days at 5°C in a refrigerator to undergo gradual water stress. Antioxidants were extracted from the stressed leaves according to CRYMCEPT (2005) and total antioxidant activity was determined according to Re et al. (1999). The total antioxidant activity was determined using a trolox (6-hydroxy-2,5,7,8-tetramethylchromane-2-carboxylic acid) standard curve and expressed as trolox equivalent. To express antioxidant activity per milligram of proteins, total protein content from the antioxidant extract above was determined by measuring its absorbance at 260 and 280 nm in the quartz cuvette. To exclude nucleic acids in the extract, total protein content at 280 nm was calculated based on Christian and Warburg equation (Caprette, 1995) as follows:
| P | = | Total water-soluble protein content (mg mL-1) |
| A280 | = | Water-soluble protein absorbance at 280 nm |
| A260 | = | Nucleic acid absorbance at 260 nm |
Data analysis: Data analysis was performed using SPSS 15.0 (SPSSR, 2006). Percentage data were arcsine-transformed to normalise their distribution. Data were subjected to one-way parametric and non parametric ANOVA based on F-test and Kruskal-Wallis test, respectively (Zar, 1997). Multiple means comparison was carried out based on Tukey honest significant difference (Tukey-HSD) test (p<0.05).
RESULTS
Plant tolerance to black sigatoka disease and water stress: The off-type banana plants were more (p<0.05) tolerant to black sigatoka disease than the MP and CP derived plants (Table 1). Moreover, the black sigatoka infection index of the MP derived plants was significantly higher (p<0.05) than that of the CP derived plants (Table 1). On the other hand, the off-type plants suffered more (p<0.05) from the water stress with higher (p<0.05) decrease in pseudostem diameter and increase in leaf senescence. The off-type plants also produced suckers during the drought stress, while none of the true-to-type plants did so during the same water stress.
Cellular mechanisms underlying plant biotic and abiotic stress responses: The off-type plants had a bigger (p<0.05) number of stomata on the leaf upper surface than the MP and CP derived plants (Table 2). They also had higher (p<0.05) stomatal density on the lower leaf surface than the MP derived banana plants. The off-type plant leaves were more (p<0.05) hydrophobic with higher (p<0.05) epicuticular wax content than the MP and CP derived plants. Water-stressed leaves of the off-type plants exhibited increased cell membrane damage with higher (p<0.05) electrolyte leakage than the true-to-type plants.
| Table 1: | Black sigatoka disease infection and drought stress responses of the off-type banana plant of landrace Uganda (±SEM) |
 | |
a, b, c: Numbers bearing the same superscript letter(s) within the column are insignificantly (p<0.05) different according to Tukey-HSD test, SEM: Standard error of the mean (n = 30) | |
| Table 2: | Stomata density, epicuticular wax content, electrolyte leakage and antioxidant activity of stressed leaves of the off-type plants (±SEM) |
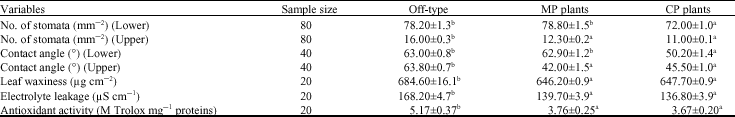 | |
a, b: Numbers bearing the same superscript letter(s) within the row are insignificantly (p<0.05) different according to Tukey-HSD test. SE: Standard error of the mean | |
The water-stressed leaves also produced more (p<0.05) antioxidants than the true-to-type plant leaves.
DISCUSSION
Plant response to black sigatoka disease and drought stress: The off-type plants were more tolerant to black sigatoka disease compared with the MP and CP derived plants. The black sigatoka disease incidence is considered to be low, mild and severe when the infection index is less than 20, 21-50 and 51-100%, respectively (Orjeda, 1998). Almost all East African highland bananas are susceptible to black sigatoka disease (Tushemereirwe, 1987) with popular landraces such as Bukoba and Embwailuma having infection indices of 24.4 and 30.5%, respectively (Msogoya et al., 2006). Tissue culture derived somaclones with increased resistance to diseases have also been reported, including off-type African plantain cv. Agbagba with mild resistance to black sigatoka (Nwauzoma et al., 2002) and off-type banana cv. Williams with resistance to yellow sigatoka disease (Trujillo and Garcia, 1996). Also reported are off-type banana plants cv. Grand nain with tolerance to panama disease (Roux, 2004), somatic embryo-derived off-type barley with tolerance to powdery mildew (Li et al., 2001) and off-type potato with tolerance to late blight (Phytophthora infestans (Mont.) de Bary) (Cassells et al., 1991). Black sigatoka is among the major diseases of banana with severe yield losses ranging from 30 to 70% (Tushemereirwe, 1987). The yield losses due to black sigatoka disease are ensued from the reduction in the number of fruits per bunch and fruit weight (Mobambo et al., 1993).
Micropropagated derived phenotypically normal plants exhibited higher susceptibility to black sigatoka disease than the CP derived plants. A reduced tolerance to fungal diseases has also been reported in many MP derived phenotypically normal plant species, including sugarcane, strawberries, African plantains and rhubarb line PC49 (Shoemark and Swartz, 1985; Péros et al., 1994; Vuylsteke, 1998; Zhao, 2004). The high susceptibility to fungal diseases of axenic in vitro regenerants has been associated with an alteration in leaf structural properties (Strange, 2003; Zhao, 2004) and a loss in plant natural defence due to removal of endophytic bacteria during tissue culture process (Hamill et al., 2005).
The off-type and MP derived banana plants manifested higher physiological injury as a result of water stress than the CP derived plants. The leaf senescence and death of the in vitro derived plants was probably due to higher water loss by transpiration as also reported in many field-grown micropropagated plant species (Marin et al., 1988; Kirdmanee et al., 1996; Herman, 2000). Water stress is among the major constraints resulting in substantial yield loss in rain fed-agriculture through a reduction in plant growth, fruit initiation and bunch weight (Robinson and Alberts, 1986; Saneoka et al., 2004). Currently, only banana groups with two B genomes such as Bluggoe (ABB) and Pisang Awak (ABB) are known to have significant natural tolerance to drought stress under tropical conditions (Simmonds, 1966; Thomas et al., 1998).
The off-type banana plants produced more suckers during water stress than the true-to-type plants. The enhanced sucker production is essential for plant survival during extensive drought stress. Earlier before this trial, it was noted that all field-grown off-type plant stools survived during the severe drought stress that hit the country in 2005, while 33% each of the MP and CP derived plant stools died. This survival mechanism possibly involves reallocation of water, nutrients and growth regulators towards the corm in the ground. The movement of nutrients, water and growth regulators from banana pseudostems towards corms after bunch harvest and their effect on the performance of next ratoon crop cycle has been documented. For instance, cutting banana pseudostems at 200 cm high above the ground after bunch harvest increases the bunch weight by 12% and reduces the time to harvest by 5% of the next ratoon crop in comparison with the removal of the whole pseudostems (Daniells and O`Farrell, 1987). More specifically, the earliness in sucker production in banana is promoted by a dense root system through its influence in cytokinin and gibberellic acid levels (Smith et al., 2001).
Cellular mechanisms underlying plant biotic and abiotic stress responses: The off-type banana plants had higher leaf stomata density, followed by the MP and CP derived plants. The observed drought stress susceptibility of the off-type and MP derived plants was probably due to higher water loss by transpiration caused by high stomatal density. A high leaf stomata density has also been reported in tissue culture derived plant species, namely bamboo, carnation and apple (Blanke and Belcher, 1989; Majada et al., 1998). The number and function of the stomata is determined at early stage of leaf development by environmental conditions, including humidity, temperatures, gaseous exchanges and liquid state of culture growth media (Jonathan et al., 1997; Majada et al., 1998). Carnation plantlets, in vitro propagated under humid condition using airtight culture vessels, had a higher stomata density, with some stomata functioning poorly when subjected to darkness, abscissic acid and polyethylene glycol (Majada et al., 1998).
The off-type banana plant leaves were more hydrophobic, which agreed with the gravimetrically determined leaf epicuticular wax content. The high leaf waxiness of the off-type plants agreed with their tolerance to black sigatoka disease. The water-dependant ascospores are the most abundant and infective structures of Mycosphaerella fijiensis (Stover and Simmonds, 1987; Stansbury et al., 2000). After the dispersal, the ascospores germinate on the moist lower leaf surface and the hyphae eventually emerge from a stoma where they either develop into conidiophores or grow across the surface to infect adjacent stomata (Stover, 1980; Stover and Simmonds, 1987). Leaf epicuticular waxiness through its influence on surface wettability has been associated with plant tolerance to fungal diseases (Gosowski, 1990; Strange, 2003). Conversely, the increased leaf waxiness of the off-type banana plants contradicted with its high water stress injury. The lack of positive correlation of leaf waxiness with drought stress tolerance has also been reported in some plant species, including maize (Ristic and Jenks, 2002). Leaf epicuticular wax deposition is influenced by environmental conditions and increases as in vitro plants acclimatise themselves to the ex vitro environment. The induction of wax deposition on leaves to protect plants against existing and subsequent water-limiting environments has also been reported in several plant species, including rose, cotton and sesame (Jenks and Ashworth, 1999; Jenks et al., 2001; Cameron et al., 2006; Kim et al., 2006; Koch et al., 2006).
The high electrolyte leakage of mannitol-dehydrated off-type plant leaves indicates membrane damage and thus a high risk of cell desiccation due to water stress. These data agreed with the observed drought stress susceptibility of the off-type plants. The high electrolyte leakage of the off-type plants positively correlated with the high antioxidant activity in the water-stressed cigar leaves. A high antioxidant activity is an indicator of increased oxidative stress and agreed well with the tolerance to black sigatoka disease of the off-type plants. The metabolites from Mycosphaerella fijiensis contain phytotoxins, which accelerate the oxidation of ascorbic acid (the most abundant antioxidant in leaf chloroplast) and induce necrotic lesions on infected banana leaves (Molina and Krausz, 1989; Busogoro et al., 2004). Thus, these authors hypothesised that the pathogenicity of black sigatoka pathogen involves the oxidative damage of leaf chloroplast. The increased antioxidant activity, however, contradicts with the water stress injury observed in the off-type plants. The lack of correlation of antioxidant activity with water-stress injury is probably due to the genetic/epigenetic difference between the off-type and true-to-type plants, which possibly affects background antioxidant levels. Antioxidants have positively been associated with plant resistance to environmental stress, including water deficit (Selote and Khanna-Chopra, 2004; Bor et al., 2003).
CONCLUSION
Off-type banana plants with increased tolerance to black sigatoka disease and plant suckering as a consequence of severe drought stress can be obtained from micropropagated landrace Uganda. This valuable variation is also associated with increased plant physiological injury as a result of drought stress. The tolerance to black sigatoka disease was possibly due to increased leaf epicuticular waxiness and total antioxidant activity, while the susceptibility to drought stress was due to high stomatal density and cell membrane instability. These findings suggest that somaclonal variation could be utilised as a tool for the improvement of plant tolerance to black sigatoka disease and increased plant suckering rate during severe water stress. The close relationship between water stress response and cell membrane injury suggests that mannitol-induced tissue dehydration might be used as a technique for early detection of off-type banana plants that are vulnerable to water stress.
ACKNOWLEDGMENT
The authors are grateful to the Commonwealth Scholarship Commission of the United Kingdom for financing (Scholarship Ref. No. TZCA-2004-129) this study.
REFERENCES
- Benson, E.E., 2000. Special symposium: In vitro plant recalcitrance-Do free radicals have a role in plant tissue culture recalcitrance. In vitro Cell. Dev. Biol. Plant, 36: 163-170.
CrossRef - Blanke, M.B. and A.R. Belcher, 1989. Stomata of apple leaves cultured in vitro. Plant Cell Tissue Organ Culture, 19: 85-89.
CrossRef - Bor, M., F. Ozdemir and I. Turken, 2003. The effects of salt stress lipid peroxidation and antioxidants in leaves of sugar beet (Beta vulgaris L.) and wild beet (Beta maritima L.). Plant Sci., 164: 77-84.
Direct Link - Busogoro, J.P., J.J. Etame, G. Harelimana, G. Lognay, J. Messiaen, P. Lepoivre and P. van Custs,em, 2004. Experimental Evidence for the Action of Mycosphaerella Fijiensis Toxins on Banana Photosynthetic Apparatus. In: Banana Improvement: Cellular, Molecular, Biology and Induced Mutation, Plymouth, Jain, S.M. and R. Swennen (Eds.). Science Publishers Inc., USA, pp: 162-181.
- Cameron, K.D., M.A. Teece and L.B. Smart, 2006. Increased accumulation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco. Plant Physiol., 140: 176-183.
CrossRefDirect Link - Campos, P.S., V. Quartin, J. Ramalho and M.A. Nunes, 2003. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol., 160: 283-292.
Direct Link - Carver, B.F. and B.B. Johnson, 1989. Partitioning of variation derived from tissue culture of winter wheat. Theor. Applied Genet., 78: 405-410.
CrossRef - Cassells, A.C., M.L. Deadman, C.A. Brown and E. Griffin, 1991. Field resistance to late blight (Phytophthora infestans Mont. De Baary) in potato (Solanum tuberosum L.) somaclones associated with instability and pleiotropic effects. Euphytica, 56: 75-80.
CrossRef - Cassells, A.C. and R.F. Curry, 2001. Oxidative stress and physiological, epigenetic and genetic variability in plant tissue culture: Implications for micropropagators and genetic engineers. Plant Cell Tissue Organ Culture, 64: 145-157.
Direct Link - Damasco, O.P, M.K. Smith, I.D. Godwin, S.W. Adkins, R.M. Smillie and S.E. Hetherington, 1997. Micropropagated dwarf off-type Cavendish bananas (Musa spp., AAA) show improved tolerance to suboptimal temperatures. Aust. J. Agric. Res., 48: 377-384.
CrossRefDirect Link - Daniells, J.W. and P.J. O`Farrell, 1987. Effect of cutting height of the parent pseudostem on yield and time to production of the following sucker in banana. Sci. Hort., 31: 89-94.
CrossRef - Gosowski, A., 1990. Influence of wheat leaf position on leaf rust severity. Euphytica, 48: 211-214.
CrossRef - Jenks, M.A. and E.N. Ashworth, 1999. Plant epicuticular waxes: Function, production and genetics. Hort. Rev., 23: 1-68.
CrossRef - Jenks, M.A., L. Andersen, R.S. Teusink and M.H. Williams, 2001. Leaf cuticular waxes of potted rose cultivars as affected by plant development, drought and paclobutrazol treatments. Physiologia Plantarum, 112: 62-70.
CrossRefDirect Link - Johnston, C., J.D. Gay, H. Staines and E.E. Benson, 2006. An in vitro oxidative stress test for determining pollutant tolerance in algae. Ecol. Indicators, 6: 770-779.
Direct Link - Jonathan, D., B. Weyers and T. Lawson, 1997. Heterogeneity in stomata characteristics. Adv. Bot. Res., 26: 317-352.
CrossRef - Kim, K.S., S.H. Park and M.A. Jenks, 2007. Changes in leaf cuticular waxes of sesame (Sesamum indicum L.) plants exposed to water deficit. J. Plant Physiol., 164: 1134-1143.
CrossRefDirect Link - Koch, K., K.D. Hartmann, L. Schreiber, W. Barthlott and C. Neinhuis, 2006. Influence of air humidity during the cultivation of plants on wax chemical composition, morphology and leaf surface wettability. Environ. Exp. Bot., 56: 1-9.
CrossRefDirect Link - Krikorian, A.D., H. Irazarry, R. Goenaga, M.E. Scott and B.E.L. Lockhart, 1999. Stability in plant and bunch traits of a French-type dwarf plantain micropropagated from the floral axis tip and five lateral corm tips of a single mother plant: Good news on the tissue culture and bad news on banana streak virus. Sci. Hortic., 81: 159-177.
CrossRef - Li, J.C., T.M. Choo, K.M. Ho, D.E. Falk and R. Blatt, 2001. Barley somaclones associated with high yield or resistance to powdery mildew. Euphytica, 121: 349-356.
Direct Link - Majada, J.P., M.L. Centeno, I. Feito, B. Fernandez and R. Sanchez-Tames, 1998. Stomatal and cuticular traits on carnation tissue culture under different ventilation conditions. Plant Growth Regul., 25: 113-121.
CrossRef - Mobambo, K.N., F. Gauhl, D. Vuylsteke, R. Ortiz, C. Pasberg-Gauhl and R. Swennen, 1993. Yield loss in plantain from black sigatoka leaf spot and field performance of resistant hybrids. Field Crops Res., 35: 35-42.
CrossRefDirect Link - Msogoya, T.J., A.P. Maerere and B.W. Grout, 2006. Field performance of micropropagated East African banana (Musa AAA East Africa) in the Eastern zone of Tanzania. Biotechnology, 5: 471-474.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nwauzoma, A.B., A. Tenkouano, J.H. Grouch, M. Pillay, D. Vuylsteke and L.A.D. Kalio, 2002. Yield and disease resistance of plantain (Musa spp. AAB group) somaclones in Nigeria. Euphytica, 123: 323-331.
CrossRefDirect Link - Peros, J.P., E. Bonnel, D. Roques and F. Paulet, 1994. Effect of in vitro culture on rust resistance and yield in sugarcane. Field Crops Res., 37: 113-119.
Direct Link - Prochazkova, D., R.K. Sairam, G.C. Srivastava and D.V. Singh, 2001. Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant Sci., 161: 765-771.
CrossRefDirect Link - Re, R., N. Pellegrini, A. Proteggente, A. Pannala, M. Yang and C. Rice-Evans, 1999. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biol. Med., 26: 1231-1237.
CrossRefDirect Link - Ristic, Z. and M.A. Jenks, 2002. Leaf cuticle and water loss in maize lines differing in dehydration avoidance. J. Plant Physiol., 159: 645-651.
Direct Link - Robinson, J.C. and A.J. Alberts, 1986. Growth and yield responses of banana cv. Williams to drip irrigation under drought and normal rainfall conditions in the subtropics. Sci. Hort., 30: 187-202.
Direct Link - Saneoka, H., R.E.A. Moghaieb, G.S. Premachandra and K. Fujita, 2004. Nitrogen nutrition and water stress effects on cell membrane stability and leaf water relation in Agrostis palustris hunds. Environ. Exp. Bot., 52: 131-138.
CrossRef - Selote, D.S. and R. Khanna-Chopra, 2004. Drought-induced spikelet sterility is associated with an inefficient antioxidant defence in rice panicles. Physiol. Plant., 121: 462-471.
Direct Link - Shoemark, P.N. and H.J. Swartz, 1985. Cultivar dependent variation in pathogen resistance due to tissue culture propagation of strawberries. HortScience, 20: 253-254.
Direct Link - Stansbury, C., S. McKirdy and G. Power 2000. Black sigatoka Mycosphaerella fijiensis: Exotic threat to Western Australia Fact Sheet.
CrossRef - Thomas, D.S., D.W. Turner and D. Eamus, 1998. Independent effects of the environment on the leaf gas exchange of three banana (Musa sp.) cultivars of different genomic constitution. Sci. Hort., 75: 41-57.
Direct Link